

Abstract
Humans pose a major threat to many species through land-use change in virtually every habitat. However, the extent of this threat is largely unknown for invertebrates due to challenges with investigating their distributions at large scales. This knowledge gap is particularly troublesome for soil macrofauna because of the critical roles many of these organisms perform as ecosystem engineers. We used a combination of high-resolution airborne Light Detection and Ranging and deep learning models to map the distribution of the ecologically important termite genus Macrotermes across a South African savanna land-use gradient, quantifying the effects of land-use change on patterns of mound densities, heights and spatial patterning. Despite significant anthropogenic alteration to landscapes, termite mounds persisted and shared a number of similarities to mounds in untransformed areas. Mean mound height was not substantially reduced in transformed landscapes, and over-dispersion of mounds at localized scales was conserved. However, mound densities were partially reduced, and height distributions in transformed areas differed to those in protected areas. Our findings suggest that mound-building termites persist even in areas of relatively high human disturbance, but also highlight important differences in termite distributions that could lead to reductions in ecosystem services provided by termites in human-modified landscapes. The persistence of at least half of mounds in human-modified landscapes could serve as starting points for savanna restoration.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Unabated human land-use change continues worldwide, leading to severe biodiversity loss and reductions in ecosystem function (Foley et al 2005, Flynn et al 2009, Newbold et al 2015). These losses increase ecosystem vulnerability and jeopardize ecosystem services and human livelihoods (Chapin III et al 2000, Potts et al 2010, Cardinale et al 2012). However, measuring changes in ecosystem function is challenging at large scales because biodiversity-ecosystem function relationships are complex and often result from intra- and/or inter-species interactions that vary across landscapes. Moreover, alterations to ecosystem function resulting from species loss can take years to manifest, exceeding the lifespan of most studies (Pringle and Tarnita 2017, Staver 2018). However, some organisms have disproportionate influence on ecosystem function, and a loss or significant disruption of these species could serve as a proxy for effects of land-use change on ecosystem function.
Globally widespread soil macrofauna such as termites, ants and earthworms play fundamental roles as ecosystem engineers. Termites are particularly important in savanna environments where their mound-building activities reorganize the physical and chemical distribution of soils throughout the landscape, creating nutrient and moisture hotspots with enhanced vegetation growth (Sileshi et al 2010, Seymour et al 2014, Davies et al 2014b). In turn, these nutrient hotspots have cascading effects on invertebrate and mammalian herbivores, and can influence vegetation and herbivory patterns over as much as 30% of savanna landscapes (Levick et al 2010b, Davies et al 2016a, 2016b). Moreover, at local scales, termite mounds are often spatially over-dispersed due to intra-specific competition between neighboring colonies, enhancing termite influence on ecosystem characteristics and function due to the even distribution of mound benefits across space (Pringle et al 2010). Termites and their mounds, as well as the spatial arrangement of colonies, can also increase agricultural yields (Evans et al 2011) and may promote ecosystem robustness to climatic change and stress events (Bonachela et al 2015, Ashton et al 2019). The persistence of keystone termite colonies and their over-dispersed spatial patterning is therefore essential for maintaining healthy, functioning savanna ecosystems.
Most studies of biodiversity loss resulting from land-use change have focused on plants and vertebrates (e.g. Pimm and Raven 2000, Ceballos and Ehrlich 2002, Flynn et al 2009). Comparatively few studies have investigated changes in invertebrate diversity, despite its wide-ranging impacts on ecosystems, and invertebrate-focused research has largely investigated local-scale effects in European and North American ecosystems (Sánchez-Bayo and Wyckhuys 2019). An urgent need exists for large-scale studies of invertebrate responses to human land-use change (Thomas et al 2019), especially studies that investigate insect population and community changes across environmental gradients. The current lack of large-scale investigations into the impacts of land-use change on invertebrates is at least partially due to difficulties in observing patterns of invertebrate richness and/or abundance over large extents, caused both by the sheer abundance of invertebrates and their small size. Mound-building termites, however, provide a rare case for assessing invertebrate responses to land-use change over large geographic areas due to the capability of remote-sensing technology to effectively map termite mounds (Levick et al 2010a, Davies et al 2014a). However, the impact of human land-use change on termites has not yet been examined at scales large enough to capture shifts in important mound characteristics such as height, which is indicative of colony size (Meyer et al 2000), density, or spatial patterning.
Effects of land-use change on termite mound distributions are also difficult to predict. Although termites have been shown to be adversely affected by human disturbance (e.g. Eggleton et al 1996, Jones et al 2003, Dosso et al 2013, Muvengwi et al 2017a), they are largely resistant to natural disturbance such as fire and herbivore activity (Davies et al 2012, Lagendijk et al 2016), and could similarly be unperturbed by human activities that do not result in whole-scale landscape changes or mound destruction, which is often the case in agricultural systems. Furthermore, human attitudes towards termites, especially in subsistence settings, can be conflicting and potentially lead to differing outcomes for termite persistence. Many people acknowledge that termites can confer some benefits to agriculture and use termite mound soil in subsistence agricultural practices (Sileshi et al 2009, Jouquet et al 2018). Humans, particularly in Africa, also consume termites as part of their diet or for medicinal purposes (Sileshi et al 2009), and could therefore be inclined to conserve termites. Conversely, however, termites are often viewed as pests that damage buildings and agricultural crops, which can drive efforts to exterminate them (Su and Scheffrahn 2000, Rouland-Lefèvre 2011, Jouquet et al 2018). Agricultural use of termite mound soil can also be destructive when cultivators disassemble mounds to distribute the enriched soil in crop fields.
Here, we investigate effects of human activity on termite mounds by combining high-resolution airborne Light Detection and Ranging (LiDAR) data with deep learning models to map distributions and heights of mounds from the widespread termite genus Macrotermes across a land-use gradient in South African savanna. We ask whether human land-use change has altered patterns in Macrotermes mound densities, heights and spatial patterning. The surveyed landscapes consisted of four land-use types, ordered here according to land-use intensity: (i) an untransformed government-managed conservation area, Kruger National Park, that has remained protected over much of the past century, (ii) a privately-owned conservation area that was converted from low density cattle farming to a protected conservation area in the 1960s, and communal areas comprised of land used either for (iii) cattle grazing and/or firewood extraction or (iv) small-scale subsistence agriculture by local communities. Termite mound locations were identified in the LiDAR data using a convolutional neural network (CNN), allowing for the rapid and consistent classification across land-use types of 47 242 termite mounds over 76 474 ha. We analyzed differences in mound density, height, and spatial patterning across the land-use types by bootstrapping equal subsamples across land-use types, and by generating and analyzing Ripley's K curves of termite mounds within each land-use type.
2. Methods
2.1. Study region
Our study took place in the Lowveld region of northeastern South Africa, a low-lying landscape dominated by savanna vegetation (figure 1). Mean annual precipitation is ∼ 630 mm yr−1 and mean annual temperature is ∼ 22° C. The dominant geology is granite, with gabbro intrusions also present (Venter et al 2003). These gabbro intrusions were excluded from the study due to the low number of Macrotermes mounds present on this geology (Davies et al 2014a). Three dominant land tenure systems are present in the region: (i) Kruger National Park (KNP), a large government-owned conservation area, (ii) Sabi-Sand Game Reserve, a privately-owned conservation area and (iii) state-owned communal areas. The area that is now KNP was officially conserved between 1898 and 1926 and today comprises almost 2 million ha. Sabi-Sand Game Reserve is a 63 000 ha privately protected area sharing an unfenced boundary (fences were removed in 1991) with KNP on its southern and eastern limits. The reserve was established in 1965 and consists of an association of freehold owners with a strong tourism-based approach to conservation. Prior to being a conservation area, the land was primarily used for low density commercial cattle farming. State-owned communal areas alongside these conservation areas are all within the Bushbuckridge municipality and are part of the former self-governing territories or 'homelands' established under the apartheid regime. Due to forced removals and relocations to the region during apartheid, population density (ranging between 150 and 300 people km−2) is high, as are levels of unemployment (Pollard et al 2003). Residents rely mostly on a combination of subsistence farming, livestock husbandry, and the consumption and trade of natural resources at informal markets (Shackleton et al 2001). Electricity is increasingly available, but firewood remains a dominant fuel for cooking, resulting in high levels of firewood extraction from communal rangelands and a subsequent reduction of woody biomass and tree cover (Wessels et al 2013). Rain-fed crops are grown around homesteads or in arable fields in close proximity to settlements.
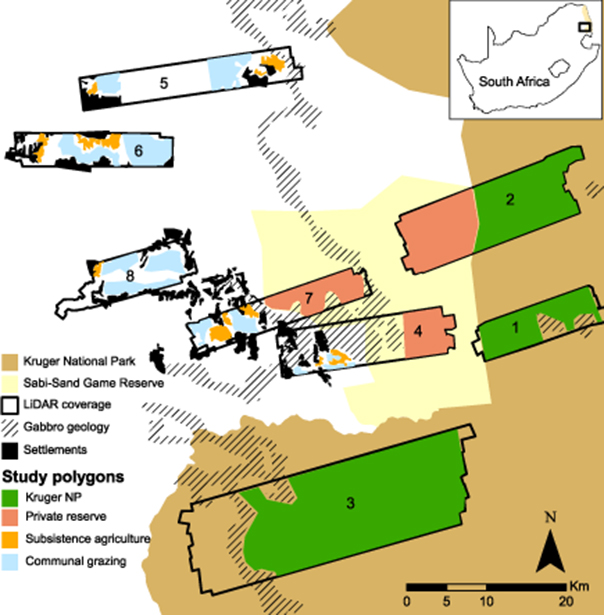
Figure 1. Study area in the Lowveld region of northeastern South Africa (see inset map) spanning (from east to west) Kruger National Park, the privately owned Sabi-Sand Game Reserve, and communal areas within the Bushbuckridge municipality. Landscapes for which airborne LiDAR data were available are delineated 1–8. Gabbro intrusions, which were excluded from analysis, are mapped within the predominantly granitic landscape. Human settlements in proximity to the study polygons are also mapped.
Download figure:
Standard image High-resolution image2.2. LiDAR surveys
Discrete-return airborne Light Detection and Ranging (LiDAR) data were collected over the study landscapes in late March/early April 2012 using the Global Airborne Observatory (GAO) Alpha system, formerly known as the Carnegie Airborne Observatory (Asner et al 2007). The LiDAR surveys were carried out at the end of the wet season when vegetation was at leaf-on stage. The aircraft was flown at approximately 2000 m above ground level, achieving an average laser shot point spacing of 2 shots m−2. Further details concerning the LiDAR specifications can be found in Asner et al (2009). LiDAR data enable high-resolution and accurate measurements of three-dimensional vegetation structure and the underlying terrain over large spatial extents by combining laser ranges from the LiDAR with location data from an embedded Global Positioning System-Inertial Measurement Unit (GPS-IMU) (Asner et al 2007). This combination results in a three dimensional 'point cloud' of known locations from each laser-surface interaction. LiDAR data points were processed to identify which laser pulses penetrated the vegetation canopy and reached the ground surface using the lasground tool packaged in LAStools software (Rapidlasso, Gilching, Germany). These points were used to interpolate a digital terrain model (DTM) of the ground surface at a 1 m resolution.
2.3. Termite mound identification
The LiDAR-derived DTM's were used to produce a hillshade map, from which 16 025 termite mounds were manually identified over 23 013 ha. Manual identification of termite mounds from hillshade maps has been shown to be accurate for mounds over 0.5 m in height (Davies et al 2014a). Two separate convolutional neural networks (CNNs) were then trained with different landscape subset sizes (256 x 256 and 512 × 512 m), and ensembled to generate a final probability. We used a CNN architecture designed for semantic segmentation, structurally similar to U-Net (Ronneberger et al 2015) but adapted for different input sizes (Brodrick et al 2019). Each contiguous patch of classified pixels was then deemed an independent mound, and the mound center was selected as the point with the maximum elevation within a 10 m radius of the cluster. Mound height was determined by taking the difference of the mound maximum elevation and the average elevation of points in a surrounding 10 m radius ring that excluded the mound itself, following Davies et al (2014a). Compared with manually identified mounds, we found precision and recall rates of 0.84 and 0.91, respectively (figure 2, appendix S1 (available online at stacks.iop.org/ERL/15/094038/mmedia)). In addition to high accuracy, the use of the CNNs facilitated a consistent treatment of mounds over large areas compared with manual identification. In total, 47 242 termite mounds were identified over 76 474 ha of study area.

Figure 2. Example hillshade images showing termite mound distributions, with and without mounds identified by the convolutional neural network (CNN) model (depicted by red circles in the right-hand panels). Hillshades were produced from airborne LiDAR-derived digital elevation models, and highlight the differences between Kruger National Park ((a)–(b), subset of Landscape 3), communal grazing land ((c)–(d), subset of Landscape 7), and agricultural fields ((e)–(f), subset of Landscape 7). See figure 1 for landscape locations.
Download figure:
Standard image High-resolution image2.4. Characterization of land-use polygons
Savanna Macrotermes in this ecosystem (southern African granitic savanna) are known to restrict mound construction to hillcrests (Levick et al 2010a, Davies et al 2014a), and woody cover can influence mound densities (Davies et al 2014a). We therefore quantified differences in topography and vegetation cover within each study polygon (see below) and land-use type. The LiDAR DTM was used to calculate the relative elevation above the nearest stream channel across each polygon by dividing the elevation above a given channel by the elevation of the nearest crest above the same channel, producing a relative elevation model (REM). Values in the REM represent proportional distances above streams with streambeds modelled as 0 and hillcrests as 1. The proportion of each polygon that consisted of stream valleys (REM between 0 and 0.05) and hillcrests (REM > 0.9) was then calculated. To calculate tree canopy height, a digital surface model (DSM) was first constructed from the LiDAR data based on interpolations of all first-return points (i.e. the top of the canopy and, where only ground returns exist, bare ground). The vertical difference between the DTM and DSM was then measured to generate a model of canopy height above ground (digital canopy model, DCM) at a 1 m resolution. The proportion of vegetation cover within each polygon was then measured as the proportion of 1 m cells that contained vegetation taller than 1 m. This height threshold ensured that only woody vegetation was included in the measurements, with herbaceous cover and bare ground excluded. We further distinguished between the proportion of tree and shrub cover in each polygon by measuring the proportion of 1 m cells that contained vegetation taller than 3 m (tree cover) and the proportion with vegetation between 1 and 3 m in height (shrub cover).
2.5. Analysis
To divide the study region into the four land-use types (subsistence agriculture, communal grazing, KNP, and private reserve), we used official boundaries for KNP and Sabi-Sand Private Reserve, and delineated Subsistence Agriculture and Communal Grazing using Google Earth images (sourced from DigitalGlobe imagery) corresponding to the 2012 LiDAR survey time period. Subsistence agriculture is clearly distinguishable from other land-uses by the presence of cleared fields, whereas communal grazing areas are undeveloped, open areas in between settlements and agricultural fields. Delineated communal grazing areas were also compared to maps in published studies (Wessels et al 2013, Mograbi et al 2015) to ensure that they covered similar extents. We excluded areas with uncharacteristic terrain (e.g. the Bushbuck Ridge in Landscape8) or unclear land-use, as well as Andover Nature Reserve in Landscape5 (see figure 1). Differences in LiDAR data availability between land-use types (table 1) resulted in varying numbers of termite mounds surveyed per land-use. To assess mound densities, we bootstrap sampled 2000, 100 ha subsets (sensitivities presented in appendix S2) within each land-use type and examined the histogram of mound densities from each set of samples. To compare distributions of mound heights between land-use types, we normalized the generated mound height histograms by the maximum binned number of mounds within the land-use type. To quantify differences in spatial patterning, we generated Ripley's K curves for each land-use type using the R package spatstat (Baddeley and Turner 2005). The Ripley' s K function is a second-order statistic that uses inter-point distances to measure deviation from spatial homogeneity. The expected distribution of features under complete spatial randomness is plotted against spatial distance and the observed distribution. When observed values fall below the expected distribution for spatial randomness, over-dispersion occurs, while values above the expected distribution indicate clustering. Because not all land-use types were contiguous (figure 1), we generated a unique Ripley's K curve for each polygon that exceeded 300 ha in size (smaller polygons were found to be dominated by boundary-condition effects and were therefore unreliable—see appendix S3). To understand the relationship between termite mound heights and densities, we used the same 2000 bootstrapped samples and compared the mean mound height to mound density within each 100 ha sample area. Finally, we examined relationships between mound density and height with woody cover for each bootstrapped sample, stratifying woody cover by the proportion within each 100 ha sample area that was under 1 m in height, between 1 and 3 m tall, and over 3 m tall.
Table 1. Termite mound, topographic and vegetation cover characteristics for each of the four land-use types.
| Land-use type | Area (ha) | No. of mounds | Bootstrapped mound density (mean mounds/ha) | Bootstrapped std. dev. of mound density (mounds/ha) | Mound density (mounds/ha) | Mean mound height (m) | Std. dev. of mound height (m) | Proportion valley (REM < 0.05) | Proportion crest (REM > 0.9) | Proportion vegetation cover > 1 m | Tree cover (proportion of vegetation cover > 3 m) | Shrub cover (proportion of vegetation cover > 1 & < 3 m) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agriculture | 2328.89 | 1039 | 0.48 | 0.10 | 0.45 | 1.19 | 0.41 | 0.09 | 0.12 | 0.11 | 0.05 | 0.06 |
| Grazing | 9341.57 | 3082 | 0.34 | 0.14 | 0.33 | 1.15 | 0.39 | 0.09 | 0.11 | 0.31 | 0.12 | 0.20 |
| Kruger | 51 196.92 | 31 994 | 0.62 | 0.27 | 0.62 | 1.27 | 0.45 | 0.11 | 0.08 | 0.27 | 0.14 | 0.13 |
| Private | 13 606.57 | 11 127 | 0.78 | 0.30 | 0.82 | 1.37 | 0.50 | 0.11 | 0.10 | 0.31 | 0.16 | 0.16 |
3. Results
Landscape topography (proportion of each polygon that was either a valley or hillcrest) did not differ substantially between study polygons or land-use types (table 1). Agricultural areas had significantly lower vegetation cover, including lower tree and shrub cover, compared with the other land-use types, which did not differ significantly from one another (table 1).
Mound densities were lowest in the communal grazing areas (mean = 0.33 mounds ha−1), followed by agricultural fields (0.45 mounds ha−1), Kruger National Park (0.62 mounds ha−1), and the private reserve (0.82 mounds ha−1) (table 1, figure 3(a)). Mound densities varied significantly among all land-use types (Kolmogorov–Smirnov (K-S) test p < 0.001 for all comparisons). Mean mound height was similar across land-use types (table 1, figure 3(b)), but the distribution of mound heights was significantly homogenized in the human-dominated landscapes. There was a substantially higher proportion of taller mounds in the protected areas, with the tallest mounds found within the private conservation areas (K-S test p < 0.001 for all comparisons, figure 3(b)). Whereas Ripley's K analyses revealed that mounds were over-dispersed at local scales in all land-use types, landscape clustering of mounds was evident in all landscapes other than the agricultural fields where mounds were randomly distributed at broader scales. The human-modified landscapes (agricultural fields and communal grazing areas) also displayed the largest variation in spatial patterning between study polygons, demonstrated by the variability in the degree of clustering and over-dispersion exhibited as well as in the distance over which either pattern persisted. This variability contrasted sharply with the almost identical patterning in each of the six protected landscape polygons. Over-dispersion of mounds also persisted over larger distances in the agricultural landscapes (up to ∼ 150 m) relative to all other land-use types (∼60-70 m) (figure 3(c)), i.e. mounds remained evenly spaced in these landscapes, but were further apart.

Figure 3. Distribution of termite mound heights (a) and densities (b) measured with airborne LiDAR across areas of varying land-use in northeastern South Africa. The height distributions were generated from all measured mounds per land-use, whereas density distributions were generated by sampling 2000 100 ha blocks with replacement from each land-use type. (c) Ripley's K (L transformation) function for termite mounds across the land-use types. The red dashed line represents the distribution of mounds under complete spatial randomness, whereas the black solid lines represent the observed mound distribution per land-use polygon (> 300 ha in extent) in each land-use type. Values below the red dashed line indicate over-dispersion (even spacing) while values above the dashed line indicate clustering. Private Reserve represents areas within Sabi-Sand Game Reserve.
Download figure:
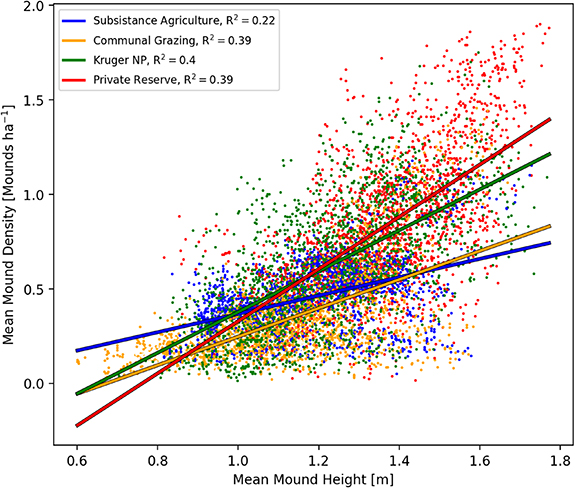
Standard image High-resolution imageAgricultural fields also displayed the weakest relationship between mound height and density across the land-use types. While there was a significant (p < 0.001) positive relationship between termite mound height and density in all land-use types, with mound height increasing linearly with density, this relationship was much stronger (evidenced by the steeper slope) within the two protected land-use types and was weakest in the agricultural fields (figure 4). Neither mound density nor height was strongly correlated with woody cover in any land-use type, nor for any woody height classification (figure S4).
{kind=link}
{kind=link}
{kind=link}
Figure 4. Termite mound heights vs densities. Points represent the mean height and density in each of the 2000 100 ha blocks from each land-use type. The lines represent linear regressions (with associated R2 values in the legend) per land-use type.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
Airborne mapping and spatial modeling revealed that despite significant anthropogenic alteration to landscapes over multiple decades, termite colonies persisted in transformed landscapes and exhibited a number of the same spatial patterning characteristics as those in protected areas. Mean termite mound height was not substantially reduced in transformed landscapes (figure 3(b)), and over-dispersion of mounds was present at localized scales in all land-use types (figure 3(c)). Moreover, although reduced, mound densities in the transformed areas were still 55% (grazing areas) and 77% (subsistence agriculture) of the densities recorded in Kruger National Park. This degree of termite resistance to land-use transformation is remarkable considering the scale of habitat modification that included clearing for agriculture, large-scale and unsustainable firewood extraction and active attempts by people to destroy and clear mounds (F. Nyathi pers. comm.). That mound densities in the human modified landscapes remained at or above half the densities of the protected areas suggests a continuation of some termite-mediated ecosystem functioning in these systems, and the potential use of termite mounds as starting points for savanna restoration given their ability to stabilize ecosystems (Bonachela et al 2015).
Our findings were also unexpected given that field-based studies have found termites to be highly sensitive to human disturbance (e.g. Eggleton et al 1996, Jones et al 2003, Muvengwi et al 2017a), and suggest that the mound-building termite, Macrotermes, is less susceptible to disturbance than other genera. Macrotermes are generalist feeders, consuming a wide range of dead organic matter, and are therefore likely able to switch their diet according to available feeding substrate. Other generalist feeders within the subfamily Macrotemitinae have been shown to be less sensitive to habitat perturbation compared with other groups, particularly soil-feeding termites (Davies et al 2012, Muvengwi et al 2017a). Macrotermes is also the largest, and likely dominant (Evans and Kasseney 2019), termite in these systems, and could exclude other termites from food resources, thereby monopolizing reduced resources in transformed landscapes. Whether dominance by Macrotermes leads to declines in other termite genera in transformed landscapes, with a concomitant reduction in ecosystem services provided by termites, was beyond the scope of our study and requires further investigation.
Notwithstanding their resistance to land-use change, we found important differences in termite mound characteristics between human-modified and protected areas. Mound densities were lower in transformed landscapes (figure 3(a)), consistent with findings of lower abundance for other taxonomic groups in response to human land-use change (Flynn et al 2009, Sánchez-Bayo and Wyckhuys 2019). However, in contrast to what might be expected, mound densities were lower in communal grazing areas relative to agricultural fields. Termite abundance has been shown to decline with agricultural clearing elsewhere (Jones et al 2003, Muvengwi et al 2017a), whereas uncleared grazing areas could be expected to be more similar to natural termite habitat (see also Hagan et al 2017 where grazing increased Trinervitermes mound densities). Subsistence agricultural practices in our study area did not include large-scale land clearing common in industrial farming (Foley et al 2005), and farming practices in the region often result in additional organic matter inputs to the soil through mulching, excess crop residue after harvesting, and the use of cattle dung for fertilizer, which likely provide increased food resources for termites. In contrast, grazing areas in the region are subject to large-scale, unsustainable firewood extraction (Wessels et al 2013), which likely lowers food availability for termites and could consequently reduce mound density. Future studies quantifying coarse woody debris across land-use types would help determine if firewood extraction is indeed a key mechanism of reduced mound densities. Over-grazing in these regions can also facilitate woody encroachment (Mograbi et al 2015), which lowers termite abundance (Leitner et al 2018). However, despite higher mound densities in agricultural areas relative to communal grazing areas, there were fewer mounds in both these land-use types relative to the protected areas, suggesting that both forms of land use have some negative effect on termite colony survival.
In contrast to clear differences in termite mound densities, mean mound heights were similar across land-use types (figure 3(b)). Mound height is strongly correlated with colony size in Macrotermes (Meyer et al 2000), and the similarities in mean mound height suggest that despite the presence of fewer colonies in human-modified landscapes, those that do establish are, on average, of similar size to those inside protected areas, suggesting that individual termite colonies are able to withstand high levels of human disturbance. Although it is possible that similar mean mound height across the land-use types is a legacy of previously extant colonies given that termite mounds can persist for exceptionally long periods of time (Erens et al 2015), abandoned mounds have been observed to remain for only a few years (e.g. 3–10 years in Australia, Wilson and Agnew 1992), reducing the likelihood of legacy effects driving the observed pattern. However, determining the proportion of active to inactive mounds in these landscapes is necessary to better understand effects of land-use change on termite colonies.
Despite similar means, the distribution of mound height was significantly truncated in agricultural fields and communal grazing areas, with most of the tallest mounds found in protected areas, particularly in privately conserved areas. Termite mound influence on savanna vegetation increases with mound size (Joseph et al 2013, Muvengwi et al 2017b), and so a higher proportion of tall (and consequently wide, Davies et al 2014) mounds in protected areas likely results in stronger mound-induced effects here relative to the human-modified landscapes. Taller termite mounds in protected areas could also contribute to increased mound densities because larger colonies likely produce more alates capable of initiating new colonies. The increased frequency of tall mounds in the private conservation areas relative to KNP is intriguing and could be due to land-use practices aimed at increasing wildlife sightings for tourists in the private reserves. These practices, which include an increased number of artificial water points and vegetation clearing along some roads, could increase food availability for termites through increased woody debris from vegetation clearing and increased dung availability from more mammalian herbivores attracted to the increased water availability. While similar spatial patterns, and therefore inter-mound distances, between the private reserves and KNP suggest consistent termite foraging ranges, increased food supply could enable the persistence of larger colonies in the private reserves.
Termite mound height also increased with mound density in all land-use types, suggesting that where termite colonies persist in large numbers, they also establish the largest colonies (see also Davies et al 2014a), likely facilitated by increased food resources. This pattern of increased mound density with height was, however, weaker in the agricultural fields, possibly because farmers actively clear mounds in a spatially stochastic fashion compared with indirect, semi-natural regulation of mounds in communal grazing areas where decreased food supply (due to woody biomass extraction) across the landscape leads to a mirrored, yet diminished, pattern of the natural areas. Farmers in the region also excavate termite mound soil for use as fertilizer (F. Nyathi, pers. comm.) and sometimes attempt to remove mounds where termites are seen as pests, altering mound structure and likely reducing height. Such clearing by humans could also lead to an increased number of small mounds in these areas, which may not be detected in the LiDAR data (Davies et al 2014a), possibly contributing to lower observed mound densities here.
Agricultural fields also displayed the strongest differences in mound spatial patterning across landscapes. Whereas Ripley's K analyses revealed that mounds were over-dispersed at local scales in all land-use types, suggesting strong inter-colony competition wherever termite mounds persist, clustering of mounds at the landscape scale was replaced by random patterning in agricultural areas. Landscape clustering arises from abiotic controls over mound distributions, with factors such as soil moisture content restricting mounds to drier hillcrests (Levick et al 2010a, Davies et al 2014a). While this clustering occurred in grazing areas, it was more variable than in the protected areas, again suggesting that mound density reductions in grazing areas occurred in a naturally mediated fashion that did not substantively disrupt their spatial patterning. In contrast, mound density reductions in agricultural areas likely stem from more direct and spatially random processes that are indifferent to abiotic controls (such as active mound clearing by humans), thereby disrupting landscape clustering. However, local-scale over-dispersion still persists in these agricultural landscapes, albeit at greater dispersion distances likely due to lower mound densities, demonstrating the powerful effect inter-colony competition has on mound patterning and the resistance of this natural process to anthropogenic disturbance. Topographic differences between land-use types were limited (table 1), suggesting that differences in mound spatial patterning in agricultural areas were not a result of selection for specific topographic regions for agriculture by people.
Given the disruptions to termite mound densities and spatial patterning in the human transformed landscapes, what are the likely consequences on ecosystem function? Over-dispersion of termite colonies at fine spatial scales leads to an even distribution of mound-associated benefits, such as nutrient and moisture enrichment (Pringle et al 2010). Since this over-dispersed pattern is not severely disrupted in any land-use we studied, the benefits of mounds remain well distributed where they occur, a remarkable finding given the significant alteration of the land surface by people (see figure 2). However, lower mound densities in human-transformed landscapes, despite similar mean colony size, will lead to reduced termite activity between mounds, which is also evidenced by the larger inter-mound distances (up to 150 m) here. Termite activities, such as foraging for dead organic matter and mound construction, influence multiple processes that enhance ecosystem function, e.g. water infiltration, nutrient cycling and decomposition of organic matter (as reviewed in Jouquet et al 2011). A decrease in termite activity could therefore lower agricultural production (Evans et al 2011) and slow natural ecosystem processes, leading to cyclical declines in resources such as woody biomass and tall tree abundance, resources already under pressure in these human-dominated landscapes (Wessels et al 2013, Mograbi et al 2017). Future studies examining changes in ecosystem function resulting from altered termite populations are essential for uncovering the impacts of termite declines on both natural ecosystem processes and human livelihoods, especially since it is unknown how changes in the assemblage composition of ecologically important taxa, such as termites, directly result in changes to the ecosystem functions they mediate (Kagezi et al 2011). Similarly, mound longevity and enhanced moisture and nutrient content after colony death are important avenues for future research. Our remote sensing approach could not quantify mound activity, which could vary across land-use types and thus influence the provisioning of ecosystem services if abandoned mounds disintegrate over short periods of time (Wilson and Agnew 1992).
There is an urgent need to understand global change impacts on under-studied but critically important groups such as insects, and while recent studies suggest widespread insect declines (Lister and Garcia 2018, Sánchez-Bayo and Wyckhuys 2019), more robust and large-scale data are needed before firm conclusions can be made (Thomas et al 2019). There is a particularly important need for more studies investigating insect population and community changes over large environmental and temporal gradients to better understand the impact of human land-use change on ecosystems and their function. Our results suggest that anthropogenic activities do have clear impacts on termite mounds, but also demonstrate surprising levels of resistance to land-use change. These findings highlight the need for careful study of changing insect communities, as well as the impacts these changes might have on ecosystem function, before allowing the rampant expansion of human activities in natural environments.
Acknowledgments
Penelope Mograbi is thanked for helpful comments on an earlier version of the manuscript The Andrew Mellon Foundation, the South African Council for Scientific and Industrial Research (CSIR) Strategic Research Panel and the South African Government Department of Science and Technology (DST), funded the LiDAR data collection used in this study. The Global Airborne Observatory is made possible by support provided by private foundations, visionary individuals, and Arizona State University.
Author contributions
GPA led the airborne data collection and research funding, ABD and CLP developed the research questions, ABD and PGB analyzed the data, ABD wrote the first draft of the manuscript, and all authors contributed to revisions.
Data accessibility
Data and code supporting the results are available at https://github.com/pgbrodrick/termiteResilience.