

Abstract
Seasonally dry tropical forests (SDTF) are located in regions with alternating wet and dry seasons, with dry seasons that last several months or more. By the end of the 21st century, climate models predict substantial changes in rainfall regimes across these regions, but little is known about how individuals, species, and communities in SDTF will cope with the hotter, drier conditions predicted by climate models. In this review, we explore different rainfall scenarios that may result in ecological drought in SDTF through the lens of two alternative hypotheses: 1) these forests will be sensitive to drought because they are already limited by water and close to climatic thresholds, or 2) they will be resistant/resilient to intra- and inter-annual changes in rainfall because they are adapted to predictable, seasonal drought. In our review of literature that spans microbial to ecosystem processes, a majority of the available studies suggests that increasing frequency and intensity of droughts in SDTF will likely alter species distributions and ecosystem processes. Though we conclude that SDTF will be sensitive to altered rainfall regimes, many gaps in the literature remain. Future research should focus on geographically comparative studies and well-replicated drought experiments that can provide empirical evidence to improve simulation models used to forecast SDTF responses to future climate change at coarser spatial and temporal scales.
Export citation and abstract BibTeX RIS
1. Introduction
Seasonally dry tropical forests (SDTF) once occupied vast amounts of tropical lands—up to 40% of all tropical forest—before large-scale deforestation (Murphy and Lugo 1986). Many of the unique properties of SDTF hinge on their rainfall regimes. SDTF rainfall regimes are characterized by distinct alternating wet and dry seasons, with some forests having dry seasons (defined as the number of months rainfall ≤100 mm) of six months or more. However, there is now abundant evidence from models and observations that suggests rainfall regimes in the seasonal tropics are changing, in part due to anthropogenic climate change (Greve et al 2014, Chadwick et al 2015). These alterations include increasing dry season length over Amazonia and increased variability in the magnitude, timing, and duration of rainfall (Feng et al 2013). It is currently not known how ongoing and future changes in rainfall may affect SDTF ecosystems, and whether SDTF will be robust or sensitive to these changes (Santos et al 2014).
SDTF provide a suite of ecosystem services such as water provisioning, flood control, tourism revenue, and pollination (Maass et al 2005), but to date no reviews have considered the extent to which these benefits to society may be compromised by climate change.
The goal of this review is to synthesize existing research to address the question: Are SDTF sensitive or resistant to decreases in rainfall and/or increases in rainfall variability? Climate change could affect SDTF through either lowered total rainfall amounts, or increased variability in the timing or duration of rainfall, either of which effectively results in rainfall shortages that occur during parts of the year that historically were wetter. Thus, our definition of drought includes both rainfall deficits that these forests may have experienced in the past, as well as novel droughts which fall outside any previously observed rainfall conditions. Throughout the review we consider two alternative hypotheses: 1) SDTF are sensitive to drought or relative water shortages because they already experience harsh environmental conditions and are near climatic thresholds, or 2) they are resistant because they tolerate or cope with seasonal water deficits. We define drought sensitivity as the changes in biological variables, states, processes, or species distributions that occur when rainfall is at or below the lower limits of what has been previously observed. By contrast, ecological resilience implies the ability to recover from changes in a relatively short amount of time, whereas resistance implies a lack of change in response to anomalously low, prolonged, or recurrent precipitation (Angeler and Allen 2016). Under the hypothesis of SDTF sensitivity, we would expect that if existing SDTF and their physiological and ecosystem processes are severely limited by rainfall and water availability, they may be extremely vulnerable to changes in rainfall amount, duration, or variability. By contrast, under the alternative hypothesis of resistance/resilience, as most SDTF species have evolved strategies such as leaf deciduousness to tolerate or avoid severe seasonal drought, it is possible that these strategies will result in few alterations in SDTF structure and function in response to future changes in rainfall regimes. Last, it is possible that SDTF responses to drought depend upon the process under consideration, with responses of different dynamics (e.g. decomposition, seedling recruitment) or species displaying a continuum of responses from highly vulnerable to highly resistant or resilient.
To evaluate whether SDTF are sensitive or resistant to changes in rainfall, we first define rainfall regimes and the conditions that constitute drought in highly seasonal tropical forests. Next we evaluate the existing literature to assess evidence that demographic processes, community dynamics, and ecosystem processes in SDTF are vulnerable or resistant/resilient to changes in rainfall, focusing on above- and belowground separately. We then discuss factors that might modulate responses to changing rainfall regimes, including deforestation and fragmentation, and also summarize work to incorporate SDTF into simulation models. We conclude by suggesting future work to improve our understanding of SDTF and climate change. Our review complements recent work that focuses on rainforests (Bonal et al 2016) and Amazon basin forests (Olivares et al 2015) in that we consider responses to altered rainfall regimes from microbial to ecosystem scales in highly seasonal tropical forests. Because our review is qualitative and synthetic, we relied first on our own knowledge and experience with this literature to select relevant studies. We supplemented this initial set of relevant papers with articles identified via searches of the Web-of-Science database using the search terms 'tropical forest' and 'drought'.
2. Rainfall and rainless regimes in SDTF
Rainfall within a given SDTF has a distinct seasonal distribution, and seasonal patterns of rainfall can also vary dramatically among SDTF. Thus, defining the term 'drought' in SDTF can be challenging, as it is possible that a variety of changes to rainfall timing, duration, and/or intensity may result in drought scenarios (section 2.2).
2.1. Variation in rainfall regimes among and within SDTF
In tropical latitudes, both total annual rainfall and rainy season length have large spatial variations and generally decrease from equatorial to subtropical regions (figure 1). In general, SDTF have a mean annual temperature >17 °C, between 200–2500 mm annual rainfall, and an annual ratio of potential evapotranspiration to precipitation >1 (Murphy and Lugo 1986). The most characteristic feature of SDTF is an extended dry season with a majority of the precipitation (∽80%) occurring within the wet season (Maas and Burgos 2011).

Figure 1 Map of (a) annual rainfall in mm, (b) wet season (defined as 80% of annual rainfall) length in days in equatorial tropics derived from Tropical Rainfall Measuring Mission (TRMM) data. Areas with >200 mm rainfall, <180 day wet season and >17 °C mean annual temperature are considered SDTF and are shaded with black hatching.
Download figure:
Standard image High-resolution imageDespite these general patterns, there is considerable inter- and intra-annual variability in rainfall within and among SDTF. Long-term rainfall records from four well-studied SDTF illustrate how distinctive seasonality is among sites, and underscore that the terms 'wet season' and 'dry season' can be overly simplistic (figure 2). In Santa Rosa, Costa Rica and Jabiru, Colombia (figures 2(a) and (d)) a short drier period (i.e. veranillo) typically occurs during the wet season. Guánica, Puerto Rico (figure 2(b)) tends to have a bimodal rainy season with spring and fall (i.e. hurricane season) rains, but neither the spring rains nor the summer dry periods are consistent. In the Yucatán Peninsula in Mexico (figure 2(c)), cold fronts (i.e. Nortes/frentes fríos) that occur during the dry season can bring lower temperatures and rainfall and thus there are three recognized seasons (Nortes, dry season, and wet season).

Figure 2 Annual precipitation at four Neotropical dry forest sites (a) Santa Rosa, Costa Rica, (b) Guánica, Puerto Rico, (c) Kaxil Kiuic, Mexico and (d) Jabiru, Colombia. The black line represents 5-day average rainfall from TRMM data. The red line represents in-situ measured monthly average rainfall. Tan shading represents wet season length defined as 80% total annual rainfall and MWL represents mean wet season length in days.
Download figure:
Standard image High-resolution image2.2. Drought concepts in seasonally dry forests
What does a drought mean in a SDTF? Climatologists have defined a number of indices (e.g. Palmer Drought Severity Index, Standardized Precipitation Index, etc.) to both diagnose and forecast drought based on a combination of data including precipitation, temperature, water supply and demand, and/or soil moisture (Zargar et al 2011, Vincente-Serrano et al 2015). Here we use an ecological concept of drought, which we define as a reduction in rainfall or a change in the timing or distribution of rainfall or rainless periods that has the potential to directly impact community- or ecosystem-level plant or microbial processes. We use conceptual models of a 'generic SDTF' with high contrast wet and dry seasons to differentiate and contrast periodic seasonal drought (i.e. alternating wet and dry seasons, figure 3(a)) from aspects of precipitation regimes that may be altered with climate change. Drought scenarios include a reduction in the total amount of rainfall during the wet season (figure 3(b)), which is the most common type of drought simulated in large-scale, throughfall reduction experiments. This type of drought has clearly occurred in the past in SDTF as low rainfall years, and is expected to increase in the future as one potential consequence of climate change (Dai 2013). In addition to drought defined as exceptionally low annual rainfall totals, there are a number of ways that increased variability of rainfall due to climate change may be perceived as ecological drought or water deficits to organisms. For example, shorter but more intense wet seasons (figure 3(c)) or a shift in when the rainy season occurs (figure 3(d)), may both result in altered dry season lengths with no change in total yearly rainfall. Moreover, changes in the quantity and timing of rainfall may co-occur (figure 3(e)). Last, it is possible that consecutive, multi-year events affect dry forests in additive or multiplicative ways that are different than individual drought years interspersed with non-drought years (figure 3(f)). Thus, there are at least five possible scenarios for how future drought might differ from seasonal rainfall in SDTF including: reduced rainfall during a given year (figure 3(b)), altered dry season length (figure 3(c)), altered timing of rainfall (figure 3(d)), reduced rainfall coupled with altered dry season length (figure 3(e)), or multi-year drought of sequential low rainfall years (figure 3(f)).
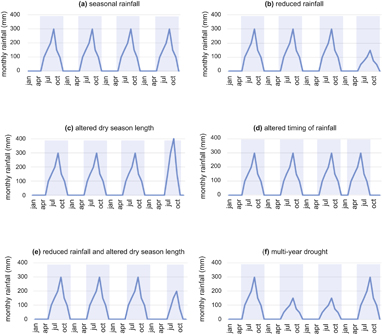
{kind=link}
{kind=link}
Figure 3 Hypothetical drought scenarios manifested as changes to monthly precipitation for four years. Blue shading defines the wet season (a) 'normal' rainfall regime with alternating wet and dry seasons, (b) annual rainfall is reduced by 50% during year 4, (c) the rainy season starts late in year 4, but annual total rainfall remains unchanged, (d) annual rainfall is equal among years, but the timing of the rainy season is shifted during year 4, (e) rainfall is reduced by 50% and the timing is shifted for year 4, (f) multi-annual drought during years 2 and 3.
Download figure:
Standard image High-resolution image{kind=link}
3. How might drought scenarios affect above- and belowground processes?
Future precipitation regimes are projected to become even more extreme in tropical regions under climate change (Malhi et al 2008, Feng et al 2013). Climate models predict decreased rainfall and increased dry periods (e.g. Maloney et al 2013, Chadwick et al 2015, Duffy et al 2015). Given the marked inter- and intra-annual variability in SDTF rainfall regimes, it is not known how alterations in the magnitude and timing of rainfall will affect the ecological dynamics of species, communities, and ecosystem processes (Vico et al 2014).
Both above- and belowground biological and ecological processes in SDTF are influenced by water availability, and the effects of increased rainfall variability, timing, or duration could alter these processes in ways that are distinct from responses to the predictable, seasonal water shortages characteristic of SDTF (figure 3(a)). Here we synthesize predictions of how this variability in rainfall regimes, including the intensity and duration of drought as well as the timing of seasonality, can all affect biological and ecological processes in SDTF at different scales. We focus on above- and belowground processes separately, because they may respond differently to drought and few studies integrate both.
3.1. Aboveground responses
Changes and variability in rainfall regimes can alter patterns of vegetative growth, physiology, and phenology. Over longer timescales, droughts may shift community dynamics and species distributions.
3.1.1. Tree phenology, physiology, and growth
In SDTF, species are distributed along a continuum of functional strategies from dense-wooded, evergreen species at one extreme to light-wooded, deciduous species at the other (Eamus 1999, Singh and Kushwaha 2005, Mendez-Alonso et al 2012). Species with high wood density have more cell wall material, low storage capacity in stems and narrow vessels, limiting hydraulic efficiency but increased resistance to drought-induced cavitation (Hacke et al 2001). These species may further resist drought by tapping subsoil water reserves with deep roots (Borchert 1994). By contrast, light-wooded species are less able to withstand xylem cavitation and are more susceptible to hydraulic failure. As a result, these species feature additional strategies to cope with drought such as high sapwood water storage, wide and conductively efficient vessels, and short-lived leaves (Ackerly 2003, Brodribb et al 2003, Meinzer et al 2008, Méndez-Alonzo et al 2012). Despite these broad generalizations, traits such as wood density do not necessarily predict leaf phenology or function at the community scale (Powers and Tiffin 2010), and there is still much to be learned about the variety of mechanisms through which species avoid, tolerate, or escape drought (Delzon 2015). Recently developed high through-put methods including osmometers to quantify turgor loss point and scanners to quantify leaf embolism are useful for identifying the potential drought response of large numbers of species in highly diverse tropical forests (Bartlett et al 2012, Delzon 2015, Maréchaux et al 2015, Brodribb et al 2016).
Many hydraulic traits are associated with drought tolerance or avoidance strategies. Leaf water potential at turgor loss point (πtlp) which is related to maintenance of cell turgor in leaves (Tyree and Jarvis 1982), and 50% loss of conductivity (Ψ50) which relates to cavitation resistance (Choat et al 2012), are useful traits for predicting species drought responses (Baltzer et al 2008, Kursar et al 2009). Species growing in dry environments have low πtlp (Bartlett et al 2012) and Ψ50 (Maherali et al 2004, Choat et al 2012), enabling them to maintain stomatal and hydraulic conductance and photosynthetic gas exchange at low soil water potentials (Sack et al 2003, Baltzer et al 2008, Kursar et al 2009). However, the narrow safety margins of SDTF species (Ψ min–Ψ50, <1 MPa, Choat et al 2012) indicate high vulnerability to drought (Choat et al 2012) and therefore future climatic scenarios with reduced rainfall may significantly impact productivity and carbon balance. Moreover, trait-dependent strategies for dealing with water limitation in SDTF may also interact with other environmental or abiotic variables. For example, shade tolerance is an important resource axis determining cavitation resistance and drought tolerance, with shade-intolerant species more vulnerable to cavitation (Markesteijn et al 2011).
Increased inter-annual rainfall variability, greater intervals between extremely wet and dry years, and particularly a decline in rainfall predicted for SDTF could influence the relative performance of species with different leaf habits and trait strategies. Decadal declines and/or increased precipitation variability may favor deciduous species (Givnish 2002, Enquist and Enquist 2011) because these species will have shorter periods of time to function without compromising their hydraulic pathway and will probably modify phenological patterns. However, predicting phenological responses requires an understanding of the relationship between phenology and traits such as stem storage capacity (Borchert 1994, Mendez-Alonzo et al 2013) and leaf age (Borchert et al 2002). In scenarios where drought is not intense enough to cause hydraulic failure but is prolonged, species with stomatal closure (isohydric) could reduce carbon uptake, resulting in carbon starvation and reduced investment in defense, and hence increased mortality by biotic agents (McDowell et al 2008, but see Sala et al 2010). Moreover, drought may affect biotic interactions between plants and other taxa (e.g. pollination, seed dispersal, frugivory, seed predation, herbivory, and soil microbes), such that climate change effects on plants ramify to other trophic levels (Parmesan and Hanley 2015). There is great potential to use remote sensing to quantify deciduousness and leaf phenology of SDTF canopies across large geographic scales and over time in relation to various climatic drivers (Cuba et al 2013). For example, Cuba et al (2013) showed that deciduousness of forest canopies in the Yucatán peninsula correlate more strongly with temperature than rainfall.
Results from throughfall exclusion (TFE) experiments, observations of natural variability among years, rainfall gradients and dendrochronology have shown that drought can impact tree growth and biomass production. To date, TFE experiments have only been performed in tropical forests with annual rainfall >2000 mm, and these studies show that rainforests are more sensitive to drought than is accounted for in models (Meir et al 2015). For example, in an Indonesian perhumid forest exposed to TFE, wood production decreased by 40% (Moser et al 2014) and in an Amazonian forest with a pronounced dry season, the decline reached 30% after seven years of TFE (da Costa et al 2010). Similarly, a water addition experiment in a seasonally moist forest in the eastern Amazon showed that stem diameter growth increased with dry season irrigation, but this effect was lagged by one year with tree growth responding to rainfall in the prior year (Vasconcelos et al 2012). Studies arrayed along natural rainfall gradients can also be a useful tool for understanding water limitation of ecosystem processes. Litterfall quantities, seasonality, and nutrient concentrations varied slightly but predictably over rainfall and successional gradients in the Yucatan peninsula (Lawrence 2005, Read and Lawrence 2003). Another study along this rainfall gradient documented systematic shifts in nutrient cycling, suggesting that nitrogen (N) limitation was strongest at lower rainfall sites while phosphorus (P) limitation increased with annual rainfall (Campo 2016). Last, dendrochronology and tree ring analysis can provide evidence of the coupling between tree growth and climate for many tropical dry forest species, as the strong seasonality causes many species to have annual growth rings (Rozendaal and Zuidema 2011). For example, a 60+ year record of seven diverse species from a dry forest in Bolivia found positive correlations between ring width and precipitation that also varied with time-scale, suggesting that most SDTF species are generally tolerant of short-term droughts, but vary in their sensitivity to multi-annual droughts (Mendivelso et al 2014).
3.1.2. Demographic processes and tree community dynamics
Evidence that drought scenarios (figure 3) may change community composition of SDTF through differential effects on demographic processes is not as well established for SDTF as it is for moist and wet tropical forest (e.g. Feeley et al 2011, Fauset et al 2012). Moreover, several factors suggest that demographic responses to drought may vary across the dry forest biome. The prevalence of recruitment modes vary substantially across SDTF, from primarily relying on establishment from seed (Vieira and Scariot 2006) to regenerating via sprouting (Swaine et al 1990, Imbert et al 1996, Dunphy et al 2000, Van Bloem et al 2003). Even within specific dry forests, both sprouting and seedling establishment vary by species, year, and type of disturbance (e.g. fire, hurricanes, clear-cutting, etc.).
A number of studies have examined seed bank and seed rain dynamics in dry forests as a function of intra- and inter-annual variation in rainfall (Castilleja 1991, Ray and Brown 1995, Martinez-Garza et al 2013, Meave et al 2012, dos Santos et al 2013). In some locations, seed bank composition differs between the wet and dry seasons (Meave et al 2012), which is not surprising if most seeds in SDTF mature or fall during the dry season (Frankie et al 1974, Martinez-Garza et al 2013). In other locations, seed fall is timed more closely to the beginning or peak of the rainy season when presumably seeds with low viability would have the best chance of establishing (Murphy et al 1995, Ray and Brown 1995, Vieira and Scariot 2006), or seed fall is comprised of species with seeds that mature at various times (Singh and Kushwaha 2006). Long dry seasons represent a bottleneck for young seedlings (Swaine et al 1990, Gerhardt 1993, Ray and Brown 1995, McLaren and McDonald 2003), suggesting that changes to dry season length will affect community composition of recruits. In addition to intra-annual variation, seedbank species richness and density in a remnant caatinga forest in Brazil varied among years and microhabitats, with significant interactions (dos Santos et al 2013). Thus, the responses of reproductive phenology to inter-annual variation in rainfall may be individualistic or under phylogenetic control, as was found in a decade-long record of reproductive phenology in a subtropical forest in China, where both flowering and fruiting were positively correlated to indices of ENSO at 2–5 month lags (Chang-Yang et al 2015). In one of the most comprehensive studies of recruitment dynamics in tropical dry forest, Maza-Villalobos et al (2013) monitored thousands of individuals 10–100 cm in height over four years in stands representing multiple successional stages in the Chamela-Cuixmala Biosphere Reserve in Mexico. Recruitment into this size class occurred primarily during the wet season, and was severely reduced following an ENSO event. By contrast, mortality rates peaked during the same period although there were complicated lag effects, which may have been caused by depleted storage reserves or ENSO-related effects on pollinator or herbivore community dynamics (Maza-Villalobos et al 2013). Marod et al (2002) suggested that the diversity of dry forest species' traits including the potential to resprout or maintain viable seedbanks, helps maintain diversity in the face of intra- and inter-annual variability in rainfall in a SDTF in Thailand. Furthermore, the reproductive phenology and productivity of dry forests appear to be highly responsive to episodic rains when rainfall regimes are unpredictable (Diaz and Granadillo 2005). However, this diversity of strategies may reduce species richness under future directional changes in rainfall regimes if only certain combinations of traits are favored, underscoring potential vulnerabilities of SDTF to changes in rainfall.
Long-term studies of community dynamics of adult trees in SDTF systems are rare, but some exist. In a study comparing two forest surveys 20 years apart in Guanacaste, Costa Rica (Enquist and Enquist 2011), extended drought conditions were accompanied by a decrease in the number of trees, mainly in the smallest sizes and in the moister habitats, as well as in the proportion of understory evergreen trees with simple leaves. A 19-year study linking tree mortality to rainfall in a 50-ha plot in Mudumulai, India (Suresh et al 2010) found mortality rates and causes varied by size class. In small size classes mortality was mostly due to fire or elephants, and mortality rates were negatively correlated to rainfall at lags of one, two, or three years. By contrast, mortality rates of trees >30 cm diameter at breast height were far lower than similarly sized trees in wet tropical forests, leading to the suggestion that large SDTF trees are resistant to inter-annual variation in climate (Suresh et al 2010). In a 10-year study in Guadeloupe, growth rates were about 50% lower and mortality increased from 1.4% to about 5% during a severe drought in 1994–95 (Imbert and Portecop 2008). By contrast, Hurricane Hugo in 1989 increased mortality to 9% and decreased growth rates by 66% (Imbert and Portecop 2008). Such long-term demographic studies are urgently needed to resolve whether and how SDTF composition will change in response to ongoing changes in climate.
3.2. Belowground responses
The effects of drought on belowground processes has received much less attention relative to aboveground dynamics. However, variability in rainfall regimes and resulting changes in soil moisture can also impact root dynamics and relationships between trees and symbionts, and ultimately these processes feedback to affect nutrient cycling and carbon storage.
3.2.1. Root dynamics
Fine roots are plants' primary organ for water and nutrient uptake, and plants can shift allocation to roots vs. shoots in order to maximize resource uptake. This suggests that fine root dynamics could be particularly sensitive to drought, either through direct effects of water deficits or indirect effects mediated by nutrient availability or other factors. In SDTF, fine root dynamics are synchronized to seasonal changes in rainfall but also respond to inter-annual precipitation anomalies (Kummerow et al 1990, Gei and Powers 2015). In addition to responding to variation in soil moisture and hence rainfall, other factors such as spatial variability in nutrients can affect fine root production and turnover (Roy and Singh 1995, Castellanos et al 2001, Powers and Peréz-Aviles 2013). Seedlings of SDTF species form deeper roots compared to species from tropical wet forests (Poorter and Markesteijn 2008), which could be a strategy to tolerate periods of soil drought. Within SDTF, root architecture may vary systematically over environmental gradients such as secondary succession or rainfall gradients. For example, a comparison of rooting depth among seedlings of 23 dry forest species showed a trade-off from vertical foraging to water storage during secondary succession, indicating that species differ in their belowground vulnerability to drought at early life stages (Paz et al 2015). Campo and Merino (2016) compared SDTF along a precipitation and forest composition gradient in the Yucatán and found increased soil carbon storage in drier sites, due to lower decomposition and higher chemical recalcitrance of fine roots.
3.2.2. Belowground symbionts and microbial community composition
Plant-soil interactions may regulate carbon cycle responses to climate change at different spatial and temporal scales (Bardgett et al 2013, Van der Putten et al 2016). Plant relationships with mycorrizhal fungi are particularly relevant for the carbon cycle given that plants transfer photosynthate carbon to fine roots where these fungi proliferate (Bardgett et al 2008, Orwin et al 2011). Plants differ in the type of mycorrhizal associations (Read et al 2004), and therefore have varied mechanisms for nutrient acquisition including the uptake of inorganic and organic forms of nitrogen (N) and phosphorus (P) (Harrison et al 2007, Bardgett et al 2008, Leigh et al 2009, Orwin et al 2011). In particular, arbuscular mycorrhizal fungi (AMF) are present in most SDTF plants (Siqueira et al 1998, Zangaro et al 2003, Mangan et al 2010), as well as ectomycorrhizal host trees (Högberg 1992, Hasselquist et al 2011), with important implications for nutrient cycling (Waring et al 2015). Both AMF (Augé 2004) and ectomycorrhizae (Lehto and Zwiazek 2011) may improve water acquisition of host plants, which could impact how SDTF trees respond to drought.
Although numerous studies report on the presence, abundance, or diversity of AMF (Allen et al 1998, Guadarrama et al 2008, Zangaro et al 2012) and ectomycorrhizae (Hasselquist et al 2011) in SDTF, very few studies have assessed how these fungi may influence the response of SDTF species to drought, or how plant-soil interactions impact the water and nutrient cycles in these forests. For instance, Gavito et al (2008) experimentally explored the effects of drought on the establishment of plant-AMF associations and found no evidence of adaptations to water stress in any of the plants or of the AMF communities. By contrast, drought limited the formation of mycorrhizal associations, although plants inoculated with AMF experienced lower water stress.
Another important plant-microbe symbiosis—symbiotic N fixation by legumes—can be affected by abiotic stresses including drought, high temperature, and salinity. Soil water limitation inhibits nodule initiation, growth and development, function, and longevity (Serraj et al 1999). Water stress also affects rhizobial survival and growth and population structure in soil (Hungria and Vargas 2000). Moreover, the regulation of N2 fixation could be altered under water stress through reduced carbon supply from the plant, oxygen permeability changes, or feedback inhibition by ureides accumulated in nodules and shoots (Valentine et al 2010). In SDTF, nodulation fluctuates seasonally (Teixeira et al 2006, González-Ruiz et al 2008, Gei and Powers 2015) and so nodules in dry forest legumes are likely to be short lived. Changes in drought intensity or in dry season length in these forests could alter nodule 'phenology' or lifespan by inducing their premature senescence. Drought stress can also delay or stop normal nodule development, as well as decrease the success of bacterial root-infection resulting in formation of ineffective nodules (Räsänen and Lindström 2003), which could be detrimental especially to seedlings. High temperatures could exacerbate these effects by decreasing survival of rhizobia, affecting the exchange of molecular signals between host plants and rhizobia, and inhibiting root-hair formation and the root-infection process (Hungria and Vargas 2000). However, mycorrhizal inoculation could alleviate the effects of drought stress and improve N2 fixation (Redecker et al 1997). It is noteworthy that the majority of studies addressing microbial responses to seasonal or exceptional drought in SDTF have focused on plant symbionts (e.g. mycorrhizae, rhizobia). However, we still have much to learn about how free-living microorganisms are affected by drought. For example, the impact of drought on soil pathogens can vary from beneficial to adverse. On the one hand, wetter conditions have been shown to be more conducive for pathogen reproduction and dispersal (Swinfield et al 2012), therefore under drier conditions there could potentially be lower numbers of soil pathogens. On the other hand, pathogens have been shown to increase the chances of mortality in drought-stressed individuals (Allen et al 2010, Spear et al 2015) through more easily infecting trees already under stress brought on by drought conditions, subsequently leading to mortality (decline spiral model; Manion 1991, Manion and Lachance 1992).
3.2.3. Ecosystem processes and nutrient cycling
Belowground ecosystem processes in tropical dry forests are sensitive to intra- and inter-annual variation in precipitation (Rohr et al 2013). For example, decomposition of leaf litter is controlled by the timing and magnitude of precipitation events (Anaya et al 2012), and annual decay rate constants increase with precipitation across SDTF (Campo and Merino 2016). Similarly, mineralization of N and P are strongly tied to rainfall patterns. Soil microbial biomass, carbon pools and the biomass C:N ratio are higher during the dry season and in drier vs. wetter sites (Singh et al 1989, Anaya et al 2007, Bejarano et al 2014). The onset of the rainy season is accompanied by a rapid increase in nutrient mineralization rates and triggers immobilization of N and P into microbial biomass (Singh et al 1989, Campo et al 1998, Austin et al 2004). In addition, the lack of rain during the dry season and during drought lowers soil respiration rates in dry tropical forests (Adachi et al 2009, Wood and Silver 2012), decoupling the positive correlation between soil temperature and respiration (Wood et al 2013).
The slower turnover of labile carbon and nutrient pools in drier forests may contribute to the negative relationship between soil carbon sequestration and mean annual precipitation across Mexican tropical dry forests (Campo and Merino 2016). At broader spatial scales, however, the relationship between soil organic carbon and aridity is hump-shaped (Delgado-Baquerizo et al 2013), suggesting that extremely dry conditions may have a negative impact on ecosystem carbon storage via decreases in carbon inputs from plant productivity or via physical processes such as erosion and fire. Similarly, the relationship between soil moisture and respiration may also be hump-shaped moving from dry to wet forests (Orchard and Cook 1983, Wood and Silver 2012), as carbon mineralization is limited by water in dry soils and oxygen in inundated soils. Therefore, the effects of shifting rainfall patterns in tropical dry forests are likely to have complex effects on belowground carbon storage, which ultimately depend on feedbacks among historical precipitation regimes, plant responses to drought, and microbial biomass growth and substrate use efficiency.
4. Regional factors that might modulate responses to drought at different timescales
Variability in rainfall regimes and drought influence both above- and belowground ecosystem processes directly. However, drought can also indirectly influence SDTF at differing timescales, and conversely, context-specific factors, other disturbances such as land-use change and forest fragmentation, and/or spatial heterogeneity—operating at both short (section 4.1) and longer timescales (section 4.2)—may modify the responses of SDTF to rainfall variability.
4.1. Short or contemporary timescales
Predictions of SDTF responses to drought scenarios (e.g. figure 3) are complicated by a number of factors that also interact with drought or modulate ecosystem responses to drought. First, elevated temperatures typically accompany drought, and both can contribute to reduced growth and/or elevated tree mortality (Allen et al 2010). Second, drought may exacerbate the effects of other disturbances, such as delayed mortality brought on by hurricanes (Van Bloem et al 2006, Imbert and Portecop 2008). Third, drought can also increase the occurrence, duration, or severity of other disturbances such as fires or insect/pathogen outbreaks; thus, drought may be the ultimate but not proximate driver of ecosystem dynamics. Similarly, landscape context (e.g. topography, distance from streams, etc.), abiotic factors such as soil fertility and depth, and distribution of forest patches in different successional stages may modify ecosystem responses to drought (Powers et al 2015), such that certain areas of a landscape are more susceptible than others to altered rainfall.
The most extensive contiguous area of SDTF can be found across South America (Miles et al 2006), the region where conservation of SDTF is also highest (accounts for 71.8% of total protected SDTF). Regions such as Africa, Southeast Asia, and Australasia, which are susceptible to anthropogenic conversion of SDTF, are extensively fragmented (Miles et al 2006). Forest fragmentation can be detrimental to these highly and uniquely diverse systems. Seed recruitment is limited across small habitat fragments, which can subsequently lead to losses in biological and genetic diversity (Nunez-Avila et al 2013).
Last, we speculate that the great climatic diversity that characterizes SDTF (figure 1), suggests that not all SDTF will respond to changes in drought in a similar fashion. For example, forests such as those in Santa Rosa, Costa Rica that experience very strong alternating wet and dry seasons with highly predictable start and end dates (figure 3(a)) may be more sensitive to changes in the timing of the wet and dry seasons (figure 3(c) and (d)) than the total amount of annual rainfall. By contrast, SDTF where rainfall is low but variable throughout the year such as in Guánica, Puerto Rico, may be more affected by the total amount of annual rainfall rather than its timing (figure 3(b)). Unfortunately, no studies we are aware of have tested the hypothesis that STDF with different rainfall regimes respond differently (or similarly) to climate change, or which mechanisms might account for this; thus, comparative studies that characterize ecological patterns and processes within and across the STDF biome remains a high priority for future research.
4.2. Decadal to evolutionary timescales
Tropical forests are dynamic and have undergone directional shifts in composition and structure over decades to millennia (Phillips and Gentry 1994, Condit 1998, Laurance et al 2004, Chave et al 2008, Feeley et al 2011). In SDTF, species composition is strongly correlated to variables related to precipitation and temperature such as water storage capacity (Santos et al 2012), water deficit (Neves et al 2015), or evapotranspiration (Saiter et al 2015). Short-term changes in species composition and forest structure have important consequences for ecosystem response to global change. Previous studies across the tropics emphasize the importance of differential species responses to drought governed by functional traits (Holbrook et al 1995, Meir and Pennington 2011). Decreasing rainfall in SDTF may favor acquisitive, drought-deciduous species as rainfall becomes insufficient to replenish deep soil water reserves (Borchert 1994, Enquist and Enquist 2011). However, decreased rainfall seasonality may favor slow-growing, evergreen species with conservative water-use strategies (Givnish 2002, Craven et al 2013).
Over evolutionary time scales, paleo-studies have shown that SDTF are strongly drought tolerant (Meir and Pennington 2011). The ecological stability of SDTF, with slow rates of compositional change and immigration rates, may constrain evolutionary responses to intensified selective pressures. Species adapted to the seasonality of SDTF show patterns of monophyly and old stem ages, suggesting persistence over evolutionary timescales (Pennington and Lavin 2015). SDTF populations in Peru and Bolivia, for example, have been stable for up to 10 million years through the drier climates of the Pleistocene. Indeed, SDTF were more extensive during cooler, drier glacial periods (Pennington et al 2009b).
Under future climate scenarios, regions such as rainforests in eastern Amazonia (Malhi et al 2008) and savanna and grasslands in India (Chaturvedi et al 2011) may become climatically suitable for SDTF, theoretically resulting in an expansion of this major biome. In order to expand into adjacent areas, SDTF species will need to disperse to track the shifting climatic envelope; however, paleo-evidence suggests that SDTF are highly dispersal limited (Pennington et al 2009b, Pennington and Lavin 2015). Moreover, the suitability of new areas for SDTF species may further depend on soil nutrient status, fire frequencies, and/or increasingly fragmented landscapes; in nutrient poor soils a transition into a savanna type ecosystem is more likely (Meir and Pennington 2011). SDTF species also lack necessary adaptations to fire (Dexter et al 2015) and moisture stress from higher temperatures or prolonged drought conditions could increase their vulnerability (Pulla et al 2015). The large rates of historic deforestation suffered by SDTF (Miles et al 2006) and current fragmented status may also affect future composition and dynamics through declines in species pools or other processes. Collectively, these factors call into question the ability of SDTF to expand to adjacent areas in geologically short periods of time. Nevertheless, many Neotropical SDTF species show wide distributions over precipitation gradients, indicating that they are climatologically more 'generalists' compared to wet forest species, which have low tolerance for dry conditions (Esquivel Muelbert et al 2016).
5. Lessons from terrestrial ecosystem models
Numerical ecosystem models can be useful tools for understanding SDTF responses to drought. Such models quantitatively synthesize numerous assumptions and hypotheses related to the real system that they represent. Model-simulated ecosystem responses to climate change or variability, including droughts, are logical consequences of underlying assumptions and hypotheses. Evaluation of model simulations therefore permits analysis of the ideas forming the ecological basis of the model. We focus here specifically on vegetation demographic models (VDMs), which are characterized by the representation and tracking of multiple (1) plant size classes, (2) plant functional types, and (3) micro-environments with a grid cell. We do not review results from big-leaf dynamic global vegetation models because such models rarely consider tree types associated with SDTF (Sitch et al 2015) and have generally not been evaluated against SDTF field data.
Holm et al (2012) carried out the first simulations of a SDTF using an individual-based VDM, ZELIG-TROP. Their simulations of Puerto Rican SDTF were generally consistent with field measurements of forest structure and basal area over the past three decades. They found that forest recovery from disturbance was highly sensitive to soil moisture, with low values of soil moisture slowing forest recovery by more than a century compared to forests that had 'ordinary' levels of soil moisture. Seiler et al (2014) used the LPJ-GUESS model to simulate tropical forests in Bolivia. Their model was able to capture the observed transition between evergreen forests and deciduous dry forests, and they identified thresholds for precipitation and water deficit beyond which leaf abscission becomes a competitive advantage. In their simulations and in satellite observations, annual GPP of SDTF was more sensitive to rainfall anomalies than that of wet tropical forests. When the LPJ-GUESS model was forced with the precipitation projections that have been projected for Bolivia at the end of the 21st century, Seiler et al (2015) found that the pure effect of decreased rainfall was a >70% reduction in SDTF vegetation carbon. Xu et al (2016) were the first to incorporate an explicit plant hydraulic scheme into model simulations focusing on SDTF. In Costa Rica and throughout Mesoamerica, they found that plant hydraulics greatly improved the model's ability to simulate responses to seasonal drought, including litterfall and phenological variation among plant functional types (PFTs). They also realistically simulated PFT-dependent responses of woody growth to inter-annual variability in rainfall. Overall, PFT responses to drought were strongly mediated by other prescribed traits including the leaf turgor loss point, xylem hydraulic conductivity, and rooting depth.
These studies suggest that SDTF would be highly sensitive to increased drought. However, work to improve model simulations of SDTF is needed. Potentially critical processes such as hydraulic failure are generally missing from models (Anderegg et al 2012). Belowground parameterizations are poorly constrained (Warren et al 2014), and implications of trade-offs between above- and belowground allocation should be explored (Doughty et al 2015). Functional diversity is clearly important for model simulations (Anderegg 2015, Xu et al 2016), but further testing of this aspect would be useful. Finally, we recommend that model development be coordinated with experimental manipulations and ongoing observational analyses.
6. Conclusions and future work
Our initial question was whether seasonally dry tropical forests, their species, and their ecological dynamics are resistant/resilient to intra- and inter-annual changes in rainfall—reflecting adaptations to predictable, seasonal drought—or whether they are sensitive, as they may already be limited by water and close to climatic thresholds. The available data, from diverse sources such as observations of seedlings, TFE experiments, dendrochronology, and modeling collectively suggest that the structure and function of SDTF will change as droughts become more frequent or severe or rainfall becomes more variable (Chadwick et al 2015, Feng et al 2013, Greve et al 2014). These changes are likely to result in reduced carbon storage as has been seen in Amazonia (Phillips et al 2009), altered biodiversity and species ranges (Enquist 2002), lowered potentials for reforestation and secondary forest regrowth (Uriarte et al 2016) and diminished capacities to provide ecosystem services. Many of these processes may display lags, such that the effects of droughts are only manifested after several years. Moreover, drier tropical forests may be slower to recover from intense or prolonged droughts (i.e. may have lower engineering resilience sensu; Angeler and Allen 2016) compared to wetter tropical forests, similar to results from a meta-analysis of drought effects on temperate zone forests that found that dry ecosystems took the longest to re-establish normal growth rates after extreme drought (Anderegg et al 2015).
Seasonally dry tropical forest ecosystems contain unique and threatened biodiversity (Pennington et al 2009a) and provide many ecosystem services that affect human well-being (Maass et al 2005). Despite our conclusion that SDTF are sensitive to changes in drought intensity, frequency, or timing, there are many remaining gaps in our knowledge. In particular, future work should establish well-replicated drought simulations or experiments, distributed across the range of dry forest climatic variation and biogeography. Comparative studies across the SDTF biome to understand which tropical dry forests and their tree species incur high mortality during droughts, and the mechanisms that underlie these responses is also a high priority, as are studies that integrate below- and aboveground responses to drought. Together, these experiments and observations should be used to improve simulation models, which in turn may be used to forecast SDTF responses to altered climates across coarser spatial scales and over a range of potential climate change scenarios.
Acknowledgments
We thank the US Department of Energy, Office of Science, Office of Biological and Environmental Research, Terrestrial Ecosystem Science (TES) Program under award number DE-SC0014363 for funding. JSP also thanks the US National Science Foundation CAREER Award DEB-1053237. This is Clemson Experimental Station technical contribution 6490. Two anonymous reviewers provided helpful comments that improved the manuscript.