
Abstract
Multicomponent mass-transport in cation-exchange membranes involves the movement of multiple species whose motion is coupled one to another. This phenomenon mediates the performance of numerous electrochemical and water purification technologies. This work presents and validates against experiment a mathematical model for multicomponent mass transport in phase-separated cation-exchange membranes (e.g., perfluorinated sulfonic-acid ionomers). Stefan-Maxwell-Onsager theory describes concentrated-solution transport. Hydrodynamic theory provides constitutive relations for the solute/solvent, solute/membrane, and solvent/membrane friction coefficients. Classical porous-medium theories scale membrane tortuosity. Electrostatic relaxation creates friction between ions. The model uses calculated ion and solvent partitioning between the external solution and the membrane from Part I of this series and incorporates the corresponding ion speciation into the transport coefficients. The proposed transport model compares favorably to properties (e.g., membrane conductivity, transference numbers, electroosmosis, and permeability) measured in dilute and concentrated aqueous binary and ternary electrolytes. The results reveal that the concentration and type of ions in the external solution alter the solvent volume fraction and viscosity in the hydrophilic pathways of the membrane, changing macroscale ionomer conductivity, permeability, and transference numbers. This work provides a physicochemical framework to predict ion-exchange-membrane performance in multicomponent systems exhibiting coupled transport.
This was Paper 2125 presented at the Atlanta, Georgia, Meeting of the Society, October 13–17, 2019. This paper is part of the JES Focus Issue on Mathematical Modeling of Electrochemical Systems at Multiple Scales in Honor of Richard Alkire.
List of symbols
| Roman | |
| Partial molar volume of species m3 mol−1 | |
| Effective molar viscous volume of species m3 mol−1 | |
| Friction coefficient between species and J s cm−5 | |
| Membrane solvent transmissibility, m4 J−1 s−1 | |
| Inverted transport coefficient between species and cm5 J−1 s−1 | |
| Molar mass of species g mol−1 | |
| Components and of matrix between species, J s cm−5 | |
| Radius of water molecule, 0.1375 nm | |
| Effective radius of hydrophilic pore accessible to hydrated ions, m | |
| Pore radius, nm | |
| Total molar solution concentration, mol dm−3 | |
| Molar concentration of species mol dm−3 | |
| Association fraction of species into species | |
| Molality of species mol kg−1solvent | |
| Moles of species mol | |
| Stoichiometric coefficient of species | |
| Transference number of species | |
| Potential of mean force on species from the membrane, J | |
| Mass-averaged velocity across the membrane, m s−1 | |
| Mass fraction of species | |
| Charge number of species | |
| Hydrodynamic friction coefficient of species in the membrane, J s cm−5 | |
| Diffusion coefficient between species and m2 s−1 | |
| Faraday's constant, 96,487 C mol−1 | |
| Domain geometric factor | |
| Ionic strength, mol m−3 | |
| Number of species | |
| Gas constant, 8.3145 J mole−1 K−1 | |
| Temperature, 298 K | |
| Inverse Debye length, m−1 | |
| membrane thickness, m | |
| Scaling constant | |
| Pressure, Pa | |
| Greek | |
| Transport coefficient of species due to chemical potential gradient of mol2 J−1 m−1 s−1 | |
| Vacuum permittivity, 8.85 × 10−12 F m−1 | |
| Relative dielectric constant | |
| Distribution factor for species in a hydrophilic domain | |
| Electrochemical potential of species relative to that of species J mol−1 | |
| (Electro)chemical potential of species J mol−1 | |
| Volume fraction of species | |
| Electric potential, V | |
| Ratio of the effective pore radius and the true pore radius | |
| Solution viscosity, Pa s | |
| Conductivity, S cm−1 | |
| Membrane water content, molwater | |
| Electroosmotic coefficient | |
| Mass density, kg m−3 | |
| Tortuosity | |
| Archie's tortuosity scaling parameter | |
| Electrostatic potential, V | |
| Electrostatic parameter | |
| Vectors and Arrays | |
| Matrix of inverted friction coefficients, cm5 J−1 s−1 | |
| Matrix of friction coefficients, J s cm−5 | |
| Flux vector of species mol m−2 s−1 | |
| Molar body force on species N mol−1 | |
| Driving force on species N m−3 | |
| Velocity vector of species m s−1 | |
| Current density, A cm-2 | |
| Matrix of driving forces, N dm−3 | |
| Matrix of association fractions | |
| Matrix of species velocities, m s−1 | |
| Subscript | |
| Infinite dilution | |
| Hydrophilic domain | |
| Reference value | |
| Superscript | |
| ' | Superficial quantity |
| Experimental Construct | |
| Referenced to membrane velocity | |
| Molecular Construct | |
| Local, microscopic quantity a hydrophilic domain | |
| Dry membrane |









































































































Transport of multiple ions in phase-separated, solvent-filled membranes dictates performance of numerous energy-storage and conversion devices.1–3 For example, recent advances in proton-exchange-membrane (PEM) fuel cells involve new catalyst alloys and cerium additives that increase kinetic performance and durability but introduce challenges related to multi-ion transport in the cation-exchange membrane.4–7 Similarly, low-temperature PEM electrosynthesis technologies have recently received intense interest, but the involved products and reactants can transport across the membrane.8–10 Membranes in redox-flow batteries (RFBs) absorb and transport numerous redox-active and supporting-electrolyte species. Often, these devices operate in aqueous environments and use perfluorinated sulfonic-acid (PFSA) ionomers as separators.2 Multicomponent transport in these materials faces the conflicting goals of promoting movement of current-carrying ions between the anode and cathode while preventing crossover of redox-active species or contaminants that decrease device performance.2,11 Although previous literature provides useful descriptions of transport in these membranes, 12–22 multi-ion transport and thermodynamics remain poorly understood.23
There are two general approaches for mass-transport in ion-exchange membranes.24,25 In the limiting regime of negligible ion and water concentrations in the membrane, dilute-solution theory is valid.24,25 Dilute-solution theory predicts that the flux of a species is proportional to the concentration gradient of plus the force an electric field applies to 24,25 This theory predicts that the number of transport coefficients (e.g., Fickian diffusion coefficients) scales by the number of mobile species in the membrane (i.e., where is the total number of species present, including mobile species and the membrane).24,25 The dilute-solution theory approach has the benefit of being relatively simple to use and understand.24,25 It also requires relatively few experiments to characterize fully all transport properties.24,25 Unfortunately, dilute-solution theory cannot describe numerous transport phenomena exhibited by ion-exchange membranes, including electroosmosis (the transport of water under an applied electric field), the flow of ions due to a water concentration gradient, and, generally, the flux of species due to a concentration gradient of species 5,23–28 As a result, measured dilute-solution diffusion coefficients in concentrated solutions are not solely properties of the material. Rather, they are effective coefficients valid only for the operating conditions for which they are measured.23







The second approach is concentrated-solution theory.24,25 This formalism is more general and is applicable to solutions that range from highly concentrated to the dilute limit, in which case it reduces to dilute-solution theory. 24,25 Concentrated-solution theory predicts that the flux of species is a linear function of the electrochemical potential gradient of all but one species present.24,25 Concentrated-solution theory is the general instantiation of classical nonequilibrium thermodynamics for multicomponent solutions.24,25,29,30 It naturally describes the coupling between forces on species and transport of species 13–15,18,24,25,29–33 The number of independent transport coefficients in concentrated-solution theory (e.g., binary diffusion coefficients) scales as as it should in real systems.25 Dilute-solution theory, therefore, contains an insufficient number of transport parameters. An unfortunate consequence of a concentrated-solution description is the introduction of a large number of parameters for multicomponent systems. For example, a typical vanadium RFB with eight species necessitates 28 different experiments to characterize the transport properties completely. Further, these transport coefficients are strong functions of concentration and membrane properties (as with dilute-solution theory) and, therefore, must be quantified across relevant conditions.13,31





In between the dilute and concentrated formalisms, there are compromise theories that incorporate certain but not all transport couplings into dilute-solution theory, such as making ion flux related to water transport or adding in electroosmosis.25,34–36 These models provide a promising method to predict multicomponent transport while remaining relatively simple.11,23,37–39 However, it is not clear under what conditions these simpler approaches are valid representations of the full concentrated-solution description.
Studies of multicomponent transport in ion-exchange membranes face a choice between using dilute-solution theories and neglecting relevant transport couplings or using concentrated-solution theory but introducing an intractable number of parameters. We reduce the intractability of concentrated-solution theory by providing a mathematical formalism to calculate transport parameters at the relevant conditions based on microscale properties of the membrane. By estimating the full set of transport properties, the model reduces the burden on extensive experimental characterization.
In this paper, we use the Stefan-Maxwell-Onsager formulation to calculate transport properties.24,25,40,41 This theory is formally equivalent to other formulations of concentrated-solution theory and attributes transport coefficients to frictional interactions between species.24,25,40,41 The frictional interactions are calculable with microscale theories.42
The paper is as follows. We outline calculation of the full matrix of Stefan-Maxwell-Onsager transport coefficients based on molecular descriptors of the system. The proposed model elucidates the mechanisms driving multicomponent transport in water-filled, cation-exchange membranes. In it, we build upon the thermodynamic model of Part I43 to calculate types, amounts, and speciation of components that move across the membrane. The theory section summarizes the pertinent concentrated-solution transport framework. It also develops a microscale-based model for transport properties and their dependence on composition and membrane material properties. The proposed model relies on physical parameters mostly available from bulk-solution measurements. In the Results and Discussion section, literature data relevant for fuel-cell and RFB membranes validate the model.
Theory
We focus on perfluorinated sulfonic-acid (PFSA) chemistry because of its extensive characterization and widespread use.2 Nafion is the most widely-used type of PFSA.2 This polymer consists of a hydrophobic polytetrafluoroethylene backbone (PTFE) with side chains that terminate in negatively charge sulfonate groups.2 Upon immersion in aqueous solutions or water vapor, the domains microphase separate into into water-filled, interconnected hydrophilic domains or "pores" and hydrophobic, PTFE structural domains.2 In this section, non-equilibrium thermodynamics is employed for multi-ion transport in the membrane structure. We provide a consistent treatment of equilibrium ionic speciation and develop a microscopic model for friction coefficients. Finally, we relate the predicted friction coefficients to macroscopic, experimentally accessible concentrated-solution transport parameters.
Multicomponent mass-transport equations
Isothermal, isotropic, multicomponent mass transport is governed by the nonequilibrium thermodynamic driving force on species balancing against the drag forces between and all other species in the system. According to the Stefan-Maxwell-Onsager theory,13,25,29





where characterizes the friction between species and and is the velocity of species The driving force for transport is29






where and are, respectively, the concentration (defined later), electrochemical potential, molar mass, and external body force on species is pressure, and is mass density. We account for electrostatic forces in the electrochemical potential rather than in a body force The membrane, species is affixed to a support (e.g., a mesh or gasket) that imparts a pinning force A force balance on the membrane dictates that this force is equal to the pressure in the membrane, 44–46 An explicit stress balance in the membrane specifies 20 Absent other external forces, substitution of Eq. 2 into Eq. 1 for each mobile species in the membrane relates the electrochemical potential gradients to species velocities13,25,29















For isothermal Gibbs–Duhem demands that


where is a superficial velocity and is the friction coefficient between species and The electrochemical potential is a function of composition, pressure, temperature, and electric state. The pressure gradient appears in Eq. 4 but not in Eq. 3 because of the pinning force on the membrane.32,45,46 In Eqs. 3 and 4, the reference velocity is that of the membrane (i.e. ). Conservation of the membrane mass provides an additional constrain that relates to the laboratory frame of reference (i.e., to the support that affixes the membrane).47 At steady state, the membrane is not actively swelling and the membrane velocity equals the velocity of the laboratory. We denote transport coefficients that depend on the frame-of-reference with a superscript of the reference species (i.e., ). If the species are chemically independent (i.e., no reactions between them; the proceeding section lifts this restriction), then for species (including all species absorbed in the membrane and the membrane), there are independent equations of this form .29 Eq. 3 in matrix form is13











where and are by 3 matrices in which the th row contains, respectively, the components of the 3D vector of driving forces and velocities of species (i.e. and where the subscript denotes a row) excluding the row and column due to linear dependence.48 The transport coefficient matrix is by where for and















Onsager reciprocal relations dictate that the friction coefficients are symmetric, 48 Consequently, there are friction coefficients. coefficients are related to binary interspecies diffusion coefficients according to25




where is the total molar concentration of the solution.

The molar concentration for a phase-separated membrane is defined either on a superficial basis (e.g., a homogenous phase) that includes the polymer volume ( where and are the moles and partial molar volume of species ), or an interstitial basis (e.g., heterogeneous phases) that only includes the electrolyte solution in the membrane pores (). We use the latter definition because it is more amenable to microscopic theories of that are derived for bulk electrolyte solutions or porous media. We neglect changes to the total molar concentration in the hydrophilic domains; the molar concentration of species is where is set to the molar concentration of salt-free water at 25 °C, (=55.2 mol dm−3). This assumption is rigorously valid for high water contents () or for membranes exchanged with cations that have molar volumes similar to water.











Transport with ion association
Many ionic species undergo ion-pair or acid-base equilibria that alter transport properties.49 Transport measurements typically control amounts of neutral components added to the system and treat the constituent ionic species as fully dissociated in solution (we call this the "Experimental Construct" and denote quantities in the construct with superscript exp). For example, sulfuric acid is treated as protons and sulfate ions. The Experimental Construct provides independent driving forces and fluxes for species. However, because friction between species depends on size and charge, microscopic models consider species in their actual, associated states (we call this the "Molecular Construct" and denote quantities in this construct with superscript ). For example, sulfuric acid is treated as protons, bisulfate, and sulfate ions.42



By accounting for how driving forces in the Molecular Construct are interdependent, Appendix A shows that the friction-coefficient matrix in the Molecular Construct, (a by matrix), is related to friction coefficients in the Experimental Construct, according to





where is a by matrix with entries and is the fraction of moles of species in the Experimental Construct, that partially associates into moles of species in the Molecular Construct












Here, and are the stoichiometric coefficients of species and respectively, in the reaction of associating with another species to form For example, protons (species ) associating with sulfate ions to form bisulfate ions (species ). Note that Theory gives the Molecular Construct transport coefficients (e.g., ) that we convert to the Experimental Construct transport coefficients (e.g., ) to calculate measured transport properties. In Part III,50 it is more convenient to make calculations using Eq. 3 in the Experimental Construct. In that case, provides 's according to the definition of For convenience, we drop the superscript outside this section for quantities in the experimental construct.















Friction coefficients
In a liquid solution consisting of solvent and ionic species in a membrane, there are six types of friction coefficients: ion/solvent, cation/anion, cation/cation, anion/anion, ion/membrane, and solvent/membrane.13,42,51,52 All but the last two types are present in bulk electrolyte solutions.51 Accordingly, we use measurements of friction coefficients in bulk solution and apply common theories for their compositional dependence to calculate their value in the hydrophilic domains of the membrane. This approach requires that the distance over which these molecular interactions occur is smaller than the size of the hydrophilic domains; this assumption is justified for the highly concentrated solutions in the membrane that strongly screen hydrodynamic influences and electrostatic interactions. A hydrodynamic model of membrane pores gives ion/membrane and solvent/membrane friction coefficients. The transport coefficients are scaled by the tortuosity and volume fraction, of the hydrophilic membrane domains to relate the transport coefficients of a single hydrophilic domain to effective, superficial membrane fluxes used in Eqs. 3 and 4.53 Tortuosity scales according to Archie's law, where is the tortuosity scaling parameter.54 is the volume fraction of the polymer backbone ( neglecting the volume of absorbed ions in the membrane), is the partial molar volume of polymer per charged group (=523.8 cm3/mole-SO3− for Nafion).2 is independent of water content and electrolyte concentration in the membrane.








Ion/solvent and ion/ion friction coefficients
For ion/solvent friction coefficients, the Stokes-Einstein equation predicts the changes with solution viscosity of the binary diffusion coefficient as a result of the drag of an ion, idealized as a sphere, moving (or rotating) through a stagnant continuum solvent42

where is the solution viscosity and the superscript denotes infinite dilution. The term in parenthesis on the right side corrects the interstitial diffusion coefficient for the tortuosity and volume fraction of hydrophilic channels. The viscosity ratio arise because the solution becomes more viscous at high ionic strengths due to increased steric interactions between ions in solution. Einstein's viscosity equation predicts how solution viscosity changes with concentration55



where is the effective molar viscous volume of species that is fit to electrolyte-solution viscosity data. Stokes–Einstein theory (Eq. 9) is widely used and generally effective at predicting the concentration dependence of ion-solvent diffusion coefficients, although agreement with experiment is imperfect.24,25,42 In particular, Stokes-Einstein theory is inaccurate for associating electrolytes,25 corrections of which are accounted for by using the Molecular Construct .


Although local viscous interactions govern ion/solvent friction, long-range electrostatics dominate ion/ion interactions.56 A "cloud" of mostly oppositely charged ions surrounds an ion in solution.56 When an external field is applied, that cloud distorts and exerts a retarding force on the ion opposing the external field.56 From this resistive force, Debye-Hückel-Onsager theory predicts that in binary electrolytes the diffusion coefficient for oppositely charged ions varies with the square-root of concentration.25,42

where is the Molecular Construct ionic strength (). Eq. 11 relates diffusion coefficients measured in bulk solution at a given ionic strength to those at other concentrations. Chapman42 and Wesselingh et al.51 suggested that, since the Debye-Hückel ion cloud is governed by the ionic strength in multicomponent electrolytes; Eq. 11 also applies to mixtures. Experiments agree with the Debye-Hückel-Onsager description that friction between similarly charged ions is negligible as they scarcely interact, or13,51



Ion and solvent/membrane friction coefficients
Debye–Hückel–Onsager theory does not apply to ionic groups attached to the polymer membrane, since they are fixed and unable to form an ionic cloud around mobile ions.51 Still, the membrane exerts a frictional force on aqueous ions and solvent from microscale-viscous interactions with the membrane walls.57 Microscale hydrodynamics predicts viscous interactions between a fluid and a solid wall.58 Species velocities and concentrations discussed up to this point are macroscopic averages and correspond to experimentally measurable quantities. In developing a microscale hydrodynamic model, we invoke microscopic, local quantities that are not experimentally accessible and are denoted with a superscript

Appendix B shows that the area-averaged, superficial velocity through the membrane (through-direction denoted as the -coordinate) where each mobile species is under an electrochemical potential gradient is




where is a hydrodynamic friction coefficient that satisfies the creeping-flow momentum balance in a pore with appropriate boundary conditions. By definition, is the sum of species velocities in the Molecular Construct weighted by their mass fractions (i.e., mass-averaged velocity, where ). In Appendix C, we demonstrate that the expression for that satisfies both the hydrodynamic prediction of Eq. 13 and frictional interactions in Eq. 3 is








where the first term on the right is due to hydrodynamic interactions directly causing friction on species and the second term is due to hydrodynamic friction on species that, in turn, exerts friction on




Following classic treatments of electrokinetics in microchannels,25 Appendix B shows that for a translationally invariant pore forming a channel with tortuosity is



where is the radius of the pore and is a function of the membrane polymer volume fraction, ,59 is dry-membrane domain spacing (2.7 nm for Nafion) and is a swelling parameter determined from microstructural characterization (1.33 for Nafion).2 is the semi-empirical geometric factor that accounts for pore shape and distribution of sizes of the hydrophilic channels, and is independent of membrane water content and ion concentration.53 Just as in Eqs. 9 and 11, the term in parenthesis on the right side corrects the interstitial hydrodynamic coefficient for the tortuosity and volume fraction of hydrophilic channels. accounts for how species distributes across the channel and equals unity when is uniformly distributed.








To establish we treat the negatively charged polymer sulfonate groups as uniformly distributed along the channel walls. Because cations are solvated, they cannot approach the walls closer than their solvated radius (i.e., the outer Helmholtz plane),60 which we set to the diameter of a water molecule nm;61 Because this study deals with high membrane hydration levels where cations are fully solvated, we do not consider cation-membrane ion-pair formation (i.e., ions complexed with the surface by dehydrating and moving to the inner Helmholtz plane).60 Consequently, ionic species are distributed across a pore of effective radius according to the linearized Poisson-Boltzmann equation.60 For this system, Appendix B shows that is given by





where is the ratio of the effective pore radius traversed by ions after accounting for solvation and the true radius is is inverse Debye length (), is bulk solvent dielectric constant (= 78.3), is vacuum permittivity, and and are modified Bessel functions of the first kind with order 0 and 1, respectively. We neglect changes in solvent concentration across the pore so that











Macroscopic transport coefficients
Equations 3 and 4 provide a microscopic description of multi-ion transport in membranes that relate species fluxes to driving forces, whereas experiments obey a macroscopic description in which experimentally controlled driving forces cause species fluxes. Fuller showed that Eq. 5 inverts to a macroscopic form32

where is the molar flux vector of species and is a component of the by symmetric matrix defined as32,53







where the membrane, species is used as a reference.

Because experimental measurements rarely ascertain the transport coefficients directly, we rewrite Eq. 17 in terms of transport coefficients that are measureable under well-defined experimental conditions, such that in terms of a controlled gradient of electrochemical potential 13,25



or, equivalently, in terms of a controlled current density

where is Faraday's constant, is the charge number of species is the transference number of species is conductivity, and is the transport coefficient between species and In the absence of concentration, pressure, or temperature gradients, for a charged species where is the electric potential. To avoid invoking an arbitrary definition of when there are concentration gradients, Eqs. 19 and 20 use the chemical potential of species relative to that of species is independent of depending only on the thermodynamic variables pressure, concentration, and temperature.25 The first terms on the right sides of Eqs. 19 and 20 specify flux due to concentration and pressure gradients and the second terms specifies transport due to migration. Because protons are present in numerous applications of cation-exchange, a convenient choice for is 13





















Equations 19 and 20 are general for isothermal transport. The transport coefficients appearing in these equations are related to the 's and are material properties of the polymer membrane that for a set composition and temperature are independent of the applied driving forces. Under certain common experimental conditions, these properties have a clear physical interpretation. Specifically, ionic conductivity, and transference numbers, relate the fluxes and current to the applied electric potential in the absence of concentration, temperature and pressure gradients





and

where the second equality provides and in terms of the 's. The electroosmotic coefficient is related to the transference number of water by the ratio which is finite even though 13 Similarly, has a straightforward physical interpretation for experiments in the absence of current; is the proportionality constant relating species fluxes under chemical potential gradients absent net ionic current () and relate to according to13










where is symmetric, which gives 's of which are independent.




The transmissibility of the membrane to water, dictates the superficial velocity of water through the membrane under an applied pressure gradient. The solvent/solvent transport coefficient, relates to measured as.13




where the membrane thickness, increases with water content from the dry thickness, for isotropic swelling 2 Equation 24 is approximate because neglects volume change on mixing of the water and membrane and neglects ionic contributions to the volume of the solution.



Parameters and Calculations
Literature reports values of and, less frequently, and for PFSA membranes. Here, we consider properties of membranes that are immersed in aqueous electrolyte solutions where membrane water content is relatively high. To calculate measured properties with the proposed model for a membrane in bulk solution at a given composition, we first calculate the water volume fraction (neglecting the volume of ions, ) and molality of ions in the membrane and the speciation of associating ions from chemical-equilibrium relations outlined in Part I.43 Although this calculation is self-consistent, model and experimental errors in electrolyte partitioning propagate to measurements and predictions of the transport properties. We relate the membrane composition to the chemical potentials of the external environment using an equilibrium model. In the steady state, we need not include a viscoelastic response of the polymer, which may be required in a transient simulation.2








Equations 6, 9, 11, and 12 give for species and excluding the membrane while Eq. 14 gives The 's specify and Eq. 7 gives Inversion of following Eq. 18 provides The components of give measured transport properties outlined in Eqs. 21–23. Matrix inversions are performed using the Python package NumPy version 1.16.











Because of the wide availability of data, we restrict our investigation to the Nafion PFSA chemistry.2,62–67 Specifically, we use data for Nafion versions N117, N115, N212, and N211. The different numbered membranes have the same molecular formulae but the N11x sequence is extruded, whereas the series N21x is cast from solution; x denotes thickness in units of mils.2 For operating parameters, we use ambient temperature (298 K) and pressure (101.3 kPa). As discussed in Supplemental Material, measurements of bulk-solution transport provide most properties at these conditions (specifically, and at a reference concentration). Parameters of ions unavailable in the literature are set to those ions of similar charge number that are available (see SM).



Table I provides the two adjusted values for the parameters of Nafion membranes. These are Archie's tortuosity scaling parameter, and the geometric transport factor, Results and Discussion show that the parameter values are the best eye-fit of calculated and measured membrane conductivity proton transference number, electroosmotic coefficient, and water-water transport coefficient. and are independent of membrane water and ion content, Results and Discussion compares model predictions with experiments.




Table I. Nafion membrane specific fitting parameters in the model.
| Parameters | Value [−] |
|---|---|
| 0.3 for data from62–64,68 | |
| 1.2 otherwise | |
| 4 |


Since membrane pretreatment and processing impact network tortuosity,2 we use two values of Archie's parameter: for the highly pretreated and conductive N117 and N115 membranes measured by Okada and co-workers reported to have a proton-form conductivity of ∼0.2 S cm−1 in liquid water 62–64,68, and for all other datasets that consistently report < 0.1 S cm−1 for proton-form membranes in liquid water at room temperature.65,69,70 Both of these values fall within the range of for a range of different types of porous media (0.3–3.4)71. for different pore shapes falls between 2 and 3, which correspond to circular- and slit-pore shape cross sections, respectively.53 This range of is lower than the value fit here. The discrepancy is likely due an extremely heterogeneous distribution of hydrophilic domain sizes that leads to a large effective 53 Porous media with parallel-type pore nonuniformities in which species transport through pores that are larger than average lead to 's that are greater than those predicted by pore shape alone.53








Results and Discussion
This section compares calculated and measured transport properties. We first consider data for membranes in dilute-aqueous solutions, partially-exchanged with proton or lithium and a mono- or multivalent cation. The external solution is sufficiently dilute so that no co-ions are present in the membrane.62–64,68 The absence of co-ions makes the measurements informative for fuel-cell membrane applications. These datasets also contain different transport coefficients that permit validation of various aspects of the model. We also consider membranes in concentrated external electrolytes that incorporate co-ions from the surrounding solution. Fewer transport measurements are available under these conditions, but they test model predictions when numerous species are present. Concentrated conditions are relevant for RFB operation.65,66 In particular, we calculate transport coefficients for membranes in aqueous solutions of sulfuric acid and vanadium sulfate that are representative of electrolytes in vanadium RFBs, which are the most studied flow-battery chemistry.72
Multicomponent transport properties of mixed-cation-form membranes in water
Okada et al.62–64,68 extensively characterized transport properties of N115 and N117 by measuring conductivity proton and lithium transference numbers electroosmotic coefficient and water transport coefficient of membranes that are partially exchanged with different cations in liquid aqueous electrolytes.




The mathematical model outlined in Part I43 calculates the water volume fraction and molality of species in the membrane (given in Figs. 4, 5 and S1 in Part I). Given these values, we calculate the transport properties of mixed-exchanged proton-alkali Nafion membranes in liquid water. Figure 1 shows measured63,73 (symbols) and calculated (lines) (a) membrane conductivity (b) proton transference number (c) electroosmotic coefficient (d) and water transport coefficient (e) ion-water transport coefficient (not measured, only calculated), and (f) ion-ion transport coefficient (not measured, only calculated) as function of the fractional proton exchange (i.e. the fraction of negatively charged polymer sulfonate group charge-balanced by protons) with various alkali metal cations. For membranes partially exchanged with alkali cations and protons, the transport coefficients are related according to and 13 Best-eye fitting of the data in Fig. 1 specifies and Figure S1 shows the same calculated transport properties as Fig. 1 for lithium-form membrane exchanged with other alkali cations ( not measured).62 The fitted and in Table I calculate transport properties for mixed lithium-alkali form membranes without adjustment.

















Figure 1. Measured63 (symbols) and calculated (lines) N11x membrane (a) conductivity (b) proton transference number (c) electroosmotic coefficient and the transport coefficients between (d) water-water (e) ion-water and (f) ion-ion in liquid water as function of fractional proton exchange, with lithium (triangles), sodium (squares), potassium (pentagons), and cesium (tilted diamonds).







Download figure:
Standard image High-resolution imageAgreement in Fig. 1 between theory and experiment is sufficient for differing membrane proton fractions and ion types. There are three cases that the model differs from experiment. The model calculates a higher sodium-exchanged membrane conductivity than does the measurement. We attribute this difference to varying experimental conditions because the measured sodium-exchanged samples have lower conductivity than the other cation-exchanged samples even when the membranes are fully in proton form (i.e. they should have identical composition), as Fig. 1a shows.63,73 The model significantly over-predicts for the lithium-exchanged membranes, as Fig. 1d shows, and for partial cesium-exchanged membranes as Fig. 1c shows. These discrepancies may be partially attributed to the high experimental uncertainty for (calculated to be ∼40% for proton-form membranes in Fig. 1d) and to lack of experimental data. Further, we assume and are independent of cation type, but cation-sulfonate interactions can alter the membrane microstructure causing disagreement between calculated and measured transport properties.74–76





As the membrane exchanges from alkali cation form to proton form, conductivity increases, plotted in Fig. 1a, because protons are much more mobile than alkali cations. Figure 1b shows that the high mobility of protons causes high except in membranes that are mostly exchanged with alkali cations (>50% exchanged). Equations 21 and 22 show that conductivity increases as Nafion exchanges from alkali ions to protons and decreases, consistent with Fig. 1c.


The high mobility of protons generates less friction for water transport through the membrane (see Eq. 6). A rising ratio thus increases as Fig. 1d confirms. In the absence of current, low-mobility alkali cations move down a gradient chemical potential of water as they are dragged by water, but a streaming potential develops to ensure electroneutrality and causes highly mobile protons to move up a water chemical potential gradient (i.e., and ).




Figure 1e shows that in fully alkali ion-exchanged membranes, protons are not available to move in the opposite direction as alkali-cations and Similarly, for fully proton-exchanged membranes. Figure 1f illustrates that similarly reaches a maximum for partially exchanged membranes and is zero for fully exchanged membranes. The values of are more than an order of magnitude greater than and which demonstrate that fluxes induced by chemical-potential gradients of ions are secondary to those induced by an equal magnitude water chemical potential gradient.






The general trends described in the preceding two paragraphs hold for all membranes exchanged with each of the alkali cations. Variations in transport properties between the different alkali ions are due to different ion-water binary diffusion coefficient at infinite dilution (given in SM), to the water volume fraction of the exchanged membrane (given in Part I43), and to the molar viscous volume of the cation-exchanged sample (given in SM). decreases with increasing alkali cation crystallographic size (i.e. Li+ > Na+ > K+ > Cs+) and has the opposite trend (i.e. Li+ < Na+ < K+ < Cs+). These different physical parameters explain the variations of transport properties in Fig. 1 for the different cation-exchanged membranes.






To explore these differences, Fig. 2 plots calculated transport properties for a 50% alkali ion-exchanged Nafion membrane (i.e. = 0.5) on contour plots for (a) conductivity, (b) proton transference number, (c) electroosmotic coefficient, and (d) water, (e) ion-water, and (f) ion-ion transport coefficients as a function of on the y-axis and on the x-axis. Each x-y point in Fig. 2 are the transport properties of Nafion partially exchanged with a hypothetical alkali ion that has a diffusion coefficient and where the membrane water volume fraction is For these calculations, we set all other properties of A+ (e.g., molar viscous volume and molar mass) to those of sodium because it is in the middle of the alkali series. To provide a reference, symbols in Fig. 2 are the and for a 50% cation-exchanged Nafion membranes in liquid water for the different alkali cations.








Figure 2. Contour plot of calculated (a) conductivity, (b) proton transference number, (c) electroosmotic coefficient, (d) and water, (e) ion-water, and (f) ion-ion transport coefficient as a function of on y-axis and on x-axis for a 50% alkali ion-exchanged Nafion membrane. Symbols plot and at 50% cation-exchanged Nafion membranes in liquid water for lithium (triangle), sodium (square), potassium (pentagon), and cesium (diamond).




Download figure:
Standard image High-resolution imageFigure 2a and 2f show that and increase with increasing cation diffusivity because more mobile ions have a higher flux for a given electric field or ion chemical-potential gradient, respectively. At low water contents, rising increases and because larger pores and lower tortuosity facilitate increased ion transport. However, at high the relation is opposite because rising decreases ion concentrations, decreasing and










This non-monotonic relationship between water content and ion-ion transport causes of partially alkali-exchanged Nafion to follow the order Li+ < Na+ < Cs+ < K+, as Fig. 1f shows. Similarly, has the order Li+ < Na+ < Cs+ < K+ because of the relationship between and as well as because lithium and sodium cause stronger viscosification of the solution in the membrane (i.e. ).





Figure 2b shows the relatively small effects and have on This explains the negligible differences in for different alkali ion-exchanged membranes seen in Fig. 1b. Figs. 2c–2e show that the water-transport properties, and all rise with increasing Higher water content increases pore size and decreases tortuosity, thereby increasing water transport.








The high value for lithium-exchanged membrane has previous been attributed to lithium "dragging" water in its large solvation shell as it transits the membrane.2,63 The effect of lithium's large solvation and resulting high friction coefficient manifests as a relatively low 61 This work shows that the low lithium diffusivity is not sufficient to explain the high value of for lithium-exchanged membranes. Rather, the large is due to the higher water content of the membrane and the resulting larger hydrophilic domains of these membranes. This finding is consistent with previous hydrodynamic models.77,78




The proposed model calculates transport properties of Nafion membranes exchanged with multivalent cations. Except for Fig. 3 shows that the transport model (lines) is in reasonable agreement with experimental measurements64 (symbols, same transport properties as Fig. 1) for a proton-form membranes exchanged with various multivalent ions as a function of membrane proton fraction (). There is relatively little difference between calculated transport properties of multivalent ion-exchanged membranes because these ions have similar and water uptake.




Figure 3. Measured64 (symbols) and calculated (lines) (a) conductivity (b) proton transference number (c) electroosmotic coefficient (Fe-exchanged membrane not measured), and the transport coefficients between (d) water-water (Fe-exchanged membrane not measured), (e) ion-water (not measured), and (f) ion-ion (not measured) of a Nafion membrane in liquid water as a function of fractional proton exchange, with calcium (right triangle), nickel (down triangle), copper (left triangle), and iron (plus sign).







Download figure:
Standard image High-resolution imageFigure 3 shows that the model over predicts water transport. Multivalent cations strongly interact with polymer sulfonate groups, altering membrane morphology through crosslinking or domain rearrangement.76 This change in polymer structure may be one source of disagreement between calculated and measured transport properties.74–76 In this case, and should be functions of ion-exchange and cation type, but the exact nature of this effect requires further investigation.


Transport in concentrated electrolytes
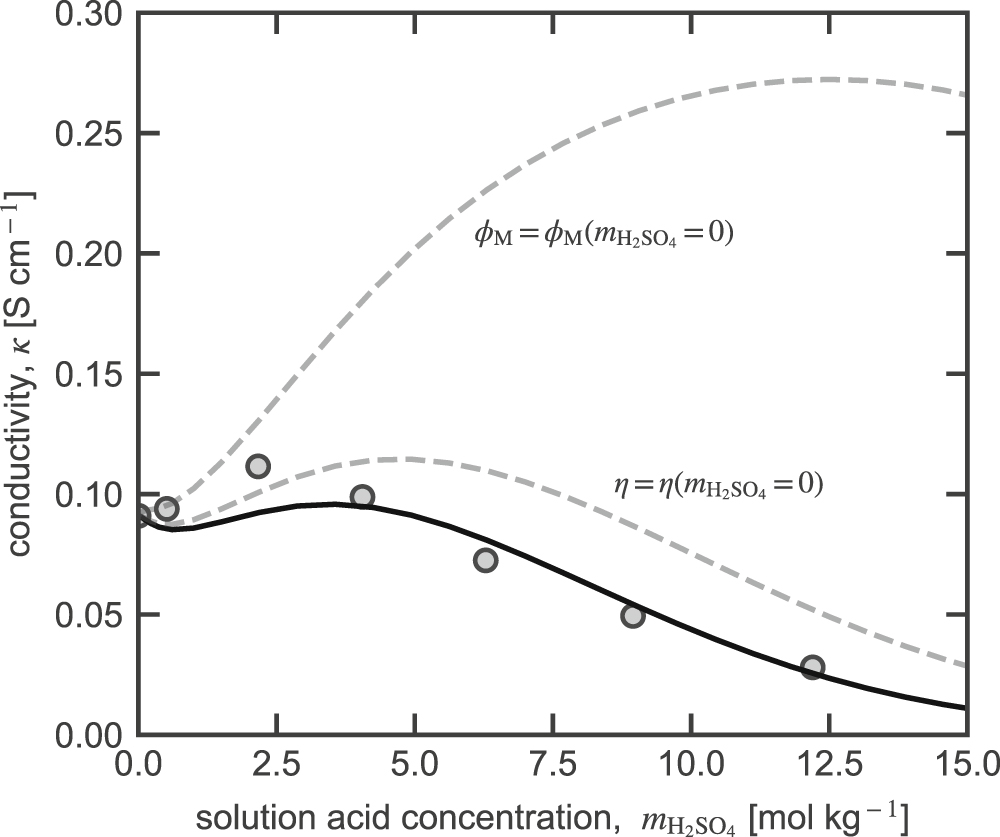
In concentrated electrolyte solutions, membrane water content and ion concentration induce large changes in transport properties. Part I43 shows that membrane water content decreases and acid uptake increases with increasing bulk electrolyte concentration. Figure 4 shows measured65,66 (circles) and calculated (solid line) N117 membrane conductivity as a function of external sulfuric acid concentration. Conductivity increases slightly up to a bulk electrolyte concentration of 4 mol kg−1. At higher concentrations, conductivity decreases with increasing electrolyte concentration.

Figure 4. Measured65 (circles) and calculated (solid lines) N117 conductivity as a function of external solution sulfuric acid concentration. Dashed lines denote model predictions with transport properties calculated if and are that of the membrane in acid-free water (i.e. or respectively).




Download figure:
Standard image High-resolution imageDashed lines in Fig. 4 show conductivity (hypothetical) if the viscosity of the electrolyte solution in the membrane or the membrane volume fraction is equal to that of the membrane in acid-free liquid water (i.e., or respectively). When viscosity of the electrolyte solution in the membrane is constant, membrane conductivity does not decrease as significantly at higher acid concentrations because proton mobility would be larger. When is held constant, the conductivity increases as the acid concentration in the membrane increases. In actuality, as bulk acid concentration increases, membrane water content decreases (see Part I43) causing increased tortuosity and decreased pore size. In agreement with Tang et al.65, the delicate balance between decreasing proton mobility and increased number of charge carriers leads to a maximum in membrane conductivity at moderate acid concentrations.





Figure 5 plots calculated (solid line) and measured67 (symbols) membrane conductivity as a function of either vanadium III, IV or V concentration with sulfuric acid such that the total sulfate concentrations is 5 mol dm-3. The conductivity is normalized to the conductivity of the membrane in vanadium-free sulfuric acid to reduce propagation of error (see Fig. 4). Based on the proposed model for intermolecular friction, there is no friction between like-charged ions (see Eq. 12). Consequently, vanadium ions and protons do not directly interact at the microscopic description of the model (i.e. ), but the presence of one still influences the other macroscopically (see Eq. 18). Although the current is carried mainly by mainly protons (dotted line shows this by plotting conductivity multiplied by proton transference number, ), as more vanadium is added to the membrane, the number of very mobile protons decreases and conductivity decreases. As Fig. 5 shows, the triply-charged V(III) displaces more protons and predicted conductivity curves are convex, whereas the singularly-charged V(V) curve is concave.



Figure 5. Measured67 (symbols) and calculated (solid lines) membrane conductivity as a function of external vanadium V(III) (pentagons), V(IV) (octagons), or V(V) (diamonds) concentration in sulfuric acid with a total sulfate concentration of 5 mol dm−3. Conductivity is normalized by the conductivity of the vanadium-free sulfuric acid solution, Dashed lines are model of the proton contribution to conductivity,


Download figure:
Standard image High-resolution imageIn addition to conductivity and proton transference number, an array of other transport properties dictate multi-ion transport in ionomer membranes. Specifically, Figure S2 shows calculated vanadium transference numbers, electroosmotic coefficients, and transport coefficients of Nafion in the same electrolytes as Fig. 5. Most of the transport properties are highly concentration dependent, and are starkly different between vanadium species. Although conductivity measurements, such as those in Fig. 5, are crucial to understand transport in these membrane, this single transport property provides a limited view of the diverse processes involved in transport.2,63,67,78 Furthermore, dilute-solution descriptions that consider only one transport parameter for each species provide an incomplete understanding of transport in many instances. As Fig. S2 demonstrates, the proposed concentrated-solution model facilitates complete specification of transport properties of multicomponent systems using only two microscale, adjustable parameters, and



Conclusions
We develop a mathematical model for multicomponent mass transport in phase-separated cation-exchange membranes based on Stefan-Maxwell-Onsager description. Microscopic theory predicts how thermodynamic and transport properties change with ion and water concentration. The model relies on two adjusted membrane-specific parameters (Archie's tortuosity parameter and pore-shape), whose values are physically reasonable and independent of water content and ion concentration. The model quantitatively agrees over wide ranges of electrolyte concentrations and compositions.
The proposed model shows that thermodynamic properties in Part I43 impact transport properties by controlling the concentration and identity of ions and water uptake. Membranes with less water have lower ion mobilities, mainly because the membrane tortuosity increases and the fraction that is conductive decreases. Moreover, increased ion concentration in the membrane increases the viscosity of the solution inside the hydrophilic domains of the membrane, further decreasing mobility. Consequently, the presence of mobile and fixed ions all impact transport both directly through Stefan-Maxwell-Onsager-type frictional interactions and indirectly by changing the structure of the membrane and the internal solution properties.
We fully specify the numerous transport coefficients involved in multicomponent transport by using concentrated-solution theory. The coefficients rigorously describe coupling of species transport. By building the model from physicochemical microscale description of transport, a paucity of experiments can specify model parameters. In Part III, we use the proposed theory to parameterize a model for multicomponent transport in a vanadium redox-flow-battery membrane, and demonstrate how concentrated-solution description in this system is essential to understand device performance.50
Acknowledgments
The work presented here was funded, in part, by Fuel Cell Performance and Durability Consortium (FC-PAD), by the Fuel Cell Technologies Office (FCTO), Office of Energy Efficiency and Renewable Energy (EERE), of the U.S. Department of Energy under contract number DEAC0205CH11231. (The information, data, or work presented herein was funded in part by an agency of the United States Government. Neither the United States Government nor any agency thereof, nor any of their employees, makes any warranty, express or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed, or represents that its use would not infringe privately owned rights. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not necessarily constitute or imply its endorsement, recommendation, or favoring by the United States Government or any agency thereof. The views and opinions of authors expressed herein do not necessarily state or reflect those of the United States Government or any agency thereof). Work by Robert Darling was supported as part of the Joint Center for Energy Storage Research, an Energy Innovation Hub funded by the U.S. Department of Energy, Office of Basic Energy Sciences under contract DE-AC02-06CH11357. The authors acknowledge fruitful discussions with Meron Tesfaye, Peter Dudenas, Ahmet Kusoglu, Douglas Kushner, Michael Gerhardt, Anamika Chowdhury, and Joseph Arthur.
Appendix A
To relate the species properties in the Experimental and Molecular Constructs, chemical equilibria of the speciation reactions give the fraction of moles of species that partially associate into moles of species 30




where the sum is over species that associate to form


The mole-weighted velocities of a fully dissociated species is the sum of the mole-weighted average velocities of its partially associated species,

where the summation is over all species that that dissociate to Equation 8 shows that Eq. A2 relates the velocity in the two constructs via



Taking the gradient of Eq. A1, multiplying the left side by and the right by and rearranging relates the driving forces in the two constructs



where the summation is over species that associate to form In matrix form, Eqs. A3 and A4 are



and

where has elements and is a full rank matrix with the number of columns greater than or equal to the number of rows, and the superscript denotes the matrix transpose. Substituting the Molecular Construct form of Eq. 5 into Eq. A5 and using Eq. A6, we have




where the power is the matrix inverse. Rearranging the Experimental Construct form of Eq. 5 and setting it equal to Eq. A7 gives


Since this equality is true for any driving force on the system, Eq. A8 rearranges to relate friction coefficients in the Experimental and Molecular Constructs


Onsager reciprocal relations holds for both and because the transformations in Eq. A9 guarantees that is symmetric since is also symmetric.




Appendix B
We consider a 2-D cylindrical channel with the -direction parallel to the channel walls and the -direction is radial. Velocity in the radial and azimuthal directions are neglected.24 Using the continuity equation, the steady-state equation of motion in the -direction for a Newtonian fluid with constant viscosity and density is24




The -velocity component is zero and where is the molar external force on species in the -direction. The average molar body force is equal to the mole-fraction weighted sum of molar body forces on each species, Radial variations of are neglected. and are the local concentration of species and fluid velocity in the -direction, respectively, with the superscript denoting a function of taking the integral average of and across the channel gives average, interstitial velocity and concentration and (i.e. and ). Note that in this section, is the interstitial velocity in a single pore. As such, is scaled by when appearing in Eqs. 3, 4, and 13. The isothermal Gibbs-Duhem (i.e., )30 equation replaces pressure changes with species (electro)chemical potential changes
























where includes the external force (i.e. includes potentials from external forces such as electrostatic field or gravity). Since the -direction is in local equilibrium, Some researchers treat electrostatic fields as an external force similar to gravity, in which case the in the Gibbs-Duhem equation includes only the chemical potential,79 whereas other researchers account for electrostatic interactions via an electrochemical potential, in which case in the Gibbs-Duhem equation is an electrochemical potential;40 Eq. B2 is the same with either approach.







We integrate Eq. B2 twice with respect to take the integral average to obtain



where

and we use the boundary conditions and requiring symmetry of the fluid velocity at the channel centerline and no-slip of the velocity at the channel walls, respectively. is the hydrodynamic friction for a single pore and is scaled by to obtain in Eq. 15 that describes macroscopic of the membrane. Comparing Eqs. 15 and B4 gives







To obtain the potential of mean force between species and the membrane dictates how is distributed across the channel relative to its average concentration, (i.e. radial distribution function of with respect to ), according to49








We treat the ions as fully solvated and that they cannot move past the Outer Helmholtz plane (i.e. for where is the effective channel radius excluding the region beyond the Outer Helmholtz plane, specifically, ). For the rest of the channel, we consider that the microscopic electrostatic potential dictates the potential of mean force such that for is referenced such that at radial position where With these expression for and linearizing B6 for small electrostatic potentials gives 56












where is the Heaviside step function ( for and for ). is the microscopic electrostatic potential that is a function of and and cannot be rigorously related the macroscopic potential 25 Poissons equation in cylindrical coordinates with constant relative permittivity dictates that for the electrostatic potential obeys56












where is the permittivity of free space and the second equality uses Eq. B7 and the definition for the inverse Debye length defined following Eq. 14. Because the microscopic electrostatic field across the channel is much greater than the electrostatic field applied across the membrane, we set the second term on the right side of Eq. B8 to zero.80 Gauss' law provides a boundary condition by dictating the total surface charge at the Outer Helmholtz plane is equal in magnitude but opposite in sign to the excess charge density in the channel,




Equations B8 and B9 dictates that The second boundary condition is symmetry of electrostatic potential at the center line, Equation B8 with these boundary conditions gives for





We find and given in Eqs. 15 and 16 using the solution for potential in Eq. B10, the distribution of ionic species in Eq. B7, and following the integration outlined in Eq. B5.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}