

Abstract
Forest characteristics, structure, and dynamics within the North American boreal region are heavily influenced by wildfire intensity, severity, and frequency. Increasing temperatures are likely to result in drier conditions and longer fire seasons, potentially leading to more intense and frequent fires. However, an increase in deciduous forest cover is also predicted across the region, potentially decreasing flammability. In this study, we use an individual tree-based forest model to test bottom-up (i.e. fuels) vs top-down (i.e. climate) controls on fire activity and project future forest and wildfire dynamics. The University of Virginia Forest Model Enhanced is an individual tree-based forest model that has been successfully updated and validated within the North American boreal zone. We updated the model to better characterize fire ignition and behavior in relation to litter and fire weather conditions, allowing for further interactions between vegetation, soils, fire, and climate. Model output following updates showed good agreement with combustion observations at individual sites within boreal Alaska and western Canada. We then applied the updated model at sites within interior Alaska and the Northwest Territories to simulate wildfire and forest response to climate change under moderate (RCP 4.5) and extreme (RCP 8.5) scenarios. Results suggest that changing climate will act to decrease biomass and increase deciduous fraction in many regions of boreal North America. These changes are accompanied by decreases in fire probability and average fire intensity, despite fuel drying, indicating a negative feedback of fuel loading on wildfire. These simulations demonstrate the importance of dynamic fuels and dynamic vegetation in predicting future forest and wildfire conditions. The vegetation and wildfire changes predicted here have implications for large-scale changes in vegetation composition, biomass, and wildfire severity across boreal North America, potentially resulting in further feedbacks to regional and even global climate and carbon cycling.
Export citation and abstract BibTeX RIS
1. Introduction
Wildfire is a dominant driver of vegetation and soil dynamics in boreal North America and interacts with forest composition, structure, regeneration, and biogeochemical cycling, as well as soil characteristics such as organic layer depth, moisture, and active layer depth (Chapin III et al 2006b, Johnstone et al 2010a). The heterogeneous landscape of boreal North America is largely driven by fire-vegetation-soil feedbacks that lead to differing forest types within the region (Bonan and Shugart 1989, Johnstone et al 2010a). Black spruce, a flammable and permafrost-tolerant species (Nikolov and Helmisaari 1992) dominates much of boreal North America, interspersed with patches of deciduous forest types and drought-tolerant, flammable Jack pine and lodgepole pine (Pinus banksiana and P. contorta, respectively) (Rogers et al 2015). Black spruce forest types tend to occur in poorly drained sites with deep organic layers, shallow active layers, and high moss growth (Johnstone et al 2010a), providing ample fuel for high severity fires which remove organic soil. Following fire, and particularly if a substantial fraction of organic soil remains, regeneration of black spruce is favored because of their semi-serotinous cones and ability to establish on deep soil organic layers (Johnstone et al 2020). The maintenance of black spruce and deep organic layers results in further buildup of organic and moss layers, resulting in a stable domain with cool, moist soils with shallow active layers and slow decomposition.
In contrast, deciduous forest types tend to occur on thin, well-drained soils with deep active layers (Chapin III et al 2006a). Litter decomposition and nutrient turnover are much faster in these forests (Melvin et al 2015), and moss growth is restricted by shading and dry soils (van Cleve and Viereck 1981). Fires in these stands maintain a stable domain of thin, dry soils and deciduous species which can readily regenerate on exposed mineral layers (Johnstone and Chapin 2006, Johnstone et al 2010a).
Fire severity, frequency, and extent in boreal North America are predicted to increase as a result of increases in lightning strike frequency, longer fire seasons, and warmer, drier conditions (Kasischke and Turetsky 2006, Kasischke et al 2010, Veraverbeke et al 2017). Severe fires consume more of the soil organic layer, allowing for potential replacement of spruce stands with deciduous species such as quaking aspen (Populus tremuloides) and birch (Betula papyrifera) (Johnstone and Chapin 2006, Beck et al 2011, Turetsky et al 2011, Johnstone et al 2020, Baltzer et al 2021). Thus, climate change may result in the breaking of this 'legacy lock' of spruce vs deciduous forest types (Johnstone et al 2010b). However, fire severity in boreal North America is strongly mediated by bottom-up controls such as fuel type and amount (Walker et al 2020b). Because deciduous stands are less flammable than spruce (Krawchuk et al 2006), negative fuel feedbacks may mediate some of the increases in fire risk that stem from top-down drivers such as climate warming (Parisien et al 2011, Rogers et al 2015). Such 'fire self-regulation' (Parks et al 2015) may limit fire ignition and progression in young post-fire stands or in stands that have converted to deciduous due to climate change. Thus, with a predicted shift towards more deciduous forests due to fire activity and climate warming (Beck et al 2011, Foster et al 2019, Mekonnen et al 2019, Hansen et al 2021), the ultimate fate of the fire regime is dependent on the interplay between increasing fuel drying and changing fuel loading, type, and flammability.
Ecological modeling provides a method of testing alternate hypotheses of potential future trajectories in ecosystem status and fire regimes (Hely et al 2009, Thonicke et al 2010, Johnstone et al 2011). The University of Virginia Forest Model Enhanced (UVAFME) is an individual tree-based forest model that has been successfully updated and tested within interior boreal Alaska (Foster et al 2019). Model outputs on forest biomass, species composition, and abiotic vegetation drivers compare well with those measured from forest inventory data and other field observations (Foster et al 2019). Previously, UVAFME predicted decreasing biomass and increasing deciduous fraction within interior Alaska as a result of climate warming and soil drying. These simulations included fire disturbance but did not include explicit links between fire weather, fuel conditions, and subsequent fire ignition and severity. To test the potential shifts in fire regimes as a result of climate change and increasing deciduous fraction, we update UVAFME to include a more mechanistic representation of fire with dynamic interactions between fuels, vegetation, fire weather, and fire activity. These updates are tested against a recent database of observed above- and belowground combustion across boreal Alaska and western Canada (Walker et al 2020a). The model is also evaluated against a suite of forest inventory data on total basal area, organic layer depth, and deciduous fraction. We then use UVAFME to simulate changing forest conditions and shifting fire probability, intensity, and severity under multiple climate change scenarios to evaluate the direction and magnitude of fire feedbacks under changing vegetation and fuel conditions. The results of these simulations provide insight into the extent to which fire self-regulation may occur under future climate change and shifting fire regimes.
2. Methods
2.1. UVAFME description and inclusion of fire processes
UVAFME simulates the establishment, growth, and mortality of individual trees on independent patches of a forested landscape. Annual tree growth, via diameter increment growth, is determined by external factors such as temperature, soil moisture, and active layer depth, as well as competition with other individuals for above- and belowground resources. Trees die through low growth or disturbances such as wildfire. Tree regeneration is mediated by climate and environmental conditions, including soil moisture, light levels, and soil organic layer depth. Climate is derived from input historical distributions of monthly temperature, precipitation, cloud cover, wind speed, and relative humidity. Daily simulations of soil moisture and freeze-thaw dynamics drive the interaction between climate, topography, forest characteristics, and soil conditions. Humus and litter decompose depending on litter type-specific characteristics (e.g. nitrogen and lignin contents) and environmental conditions such as moisture, light level, and active layer depth. For a more in-depth description of UVAFME, see Foster et al (2019) or the UVAFME GitHub page (https://uvafme.github.io).
We updated UVAFME to include shrub growth and allometry. Allometry observations from Berner et al (2015) and the Biomass and Allometry Database (Falster et al 2015) were used to create new shrub allometries to calculate annual diameter growth, shrub stem length, and biomass. We additionally updated the model to include growth limitation from inundation based on equations from Botkin (1993). See supplementary material section 4 for a more detailed description of these methods.
The fire subroutines within UVAFME now incorporate process-based representation of fire ignition and intensity based on equations from Thonicke et al (2010), Rothermel (1972), and other experimental and modeling studies describing fire ignition, intensity, and subsequent above- and below-ground combustion and vegetation mortality (van Wagner 1983, Scott and Reinhardt 2001). In this new version of UVAFME, ignition events are determined stochastically on a daily basis, based on an average site-level lightning strike frequency multiplied by a fire danger index (FDI). FDI is calculated based on the ratio of fuel moisture and fuel moisture of extinction (equation (S25) in the supplementary material available online at stacks.iop.org/ERL/17/025006/mmedia). As in Thonicke et al (2010), fuel characteristics are calculated as the weighted sum of characteristics across different fuel types. In this version, we break surface fuel types into dead needles, dead leaves, twig litter, dead and live moss, small and large branch litter, and live foliage and twigs from trees and shrubs less than 1.83 m in height. During crown fires (see below), live tree branches and crowns are also included in fuels. Organic soil and bole and root litter burn, but are not used in calculations of fire behavior. Surface fuel moisture is calculated using the Canadian Forest Fire Danger Rating System (CFFDRS) fine fuel moisture code (FFMC) (Wang et al 2015). Root, bole, and duff moisture is calculated using the CFFDRS drought moisture code (DMC).
Fire rate of spread (m min−1) and surface intensity (kW m−1) are calculated based on surface fuel loading, geometry (i.e. bulk density and surface area to volume ratio; SAV), and moisture, and wind speed. As in Rothermel (1972) and Thonicke et al (2010), this rate of spread is calculated assuming the fire has reached a quasi-steady state, with horizontally homogenous fuels. Thus, we do not calculate spatial propagation of fires to adjacent sites or grid cells, but rather calculate a potential rate of spread within the forest plot and use this variable to calculate subsequent fire behavior (e.g. intensity, combustion) on that plot. Fires with a surface intensity less than 50 kW m−1 cannot be sustained and are extinguished. Otherwise, surface fuels, boles, root litter, and humus burn based on fuel moisture and fire duration (van Wagner 1972, Thonicke et al 2010). We also include the potential for passive and active crown fires in this version of UVAFME, as per Scott and Reinhardt (2001).
Individual tree mortality is calculated based on cambial damage and percentage crown scorch (Peterson and Ryan 1986). Cambial damage is mediated by tree size and bark thickness. Smaller trees with thinner bark will have greater damage from the same fire than a larger, thicker-barked tree. Taller trees will have a lower crown scorch percentage, leading to lower mortality. See supplementary material section 4 for a more detailed description of these methods.
2.2. Study area and observational data
Our study area is defined as the boreal forest within the Core and Extended Domains of NASA's Arctic Boreal Vulnerability Experiment (ABoVE) in interior Alaska and western Canada (figure 1).
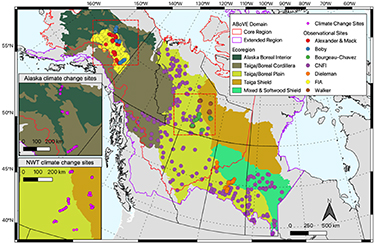
Figure 1. Study area and testing sites. Observation points are from a suite of available forest inventory and combustion field sites (table S1 in the supplementary material). Ecoregion delineations are based on the US EPA level II CEC ecoregions (US EPA 2015). Insets show the climate change application sites.
Download figure:
Standard image High-resolution imageSpecifically, we test UVAFME at sites across this domain to ensure that the model can reliably capture forest characteristics and responses to environmental conditions across a wide range of site and forest types, thereby increasing our confidence in its ability to simulate forest dynamics under novel climate and environmental conditions. Due to a lack of information on future lightning strike frequency for many of the Canadian provinces, we only test future climate and fire regimes at 300 of the combustion testing sites within Alaska and the Northwest Territories where these data are available (Veraverbeke et al 2017).
To test model updates, we compare model output to field observations from a suite of different forest inventory (n = 1026) and combustion databases (n = 335) (figure 1, table S1 in the supplementary material). Forest inventory data were chosen to span the study region and to include observations of species-specific basal area, organic layer depth, and stand age at the time of measurement. Combustion data from Walker et al (2020a) were filtered to include those with known stand age at the time of burn, day of burn, and pre- and post-fire above- and below-ground biomass and organic layer depth. See supplementary material section 2 for further details. Input climate, site, soil, and species parameters for UVAFME were derived as in Foster et al (2019). See supplementary material section 3 for further details.
2.3. Model testing
To confirm the ability of UVAFME to reliably simulate forest dynamics and conditions, we compared model output on total basal area, deciduous fraction, and organic layer depth to that measured by 1026 inventory sites across the study region. Through the Monte Carlo aggregation of hundreds of independent plots, gap models such as UVAFME are designed to represent landscape-scale forest characteristics (Shugart 1984, Bugmann et al 1996), and are thus most appropriately compared to an average of a robust sampling of replicate forest plots. Because many of the inventory sites in our compiled database only had one sampling plot, we grouped sites by Level II US EPA CEC Ecoregion (US EPA 2015) and forest type. Forest types were defined by the species or collection of species that made up at least 75% of the total basal area in the inventory data (see supplementary material section 2). At each site, UVAFME was run from bare ground initiation (each with 200 independent 500 m2 plots) until reaching a given site's stand age (obtained from inventory data). Average simulated and observed total basal area (m2 ha−1), deciduous fraction by basal area (%), and organic layer depth (cm) were compared across the ecoregion and forest type groupings. T-tests were conducted within each grouping between modeled and measured values to determine if there were any significant (p < 0.01) differences.
We additionally compared model output on above- and belowground combustion (kgC m−2) to observations from Walker et al (2020a). We again grouped these sites by Level II US EPA CEC Ecoregion, but not by forest type, as species composition was unknown for many of the combustion sites. For these simulations, we ran UVAFME from bare ground initiation (each with 200 independent 500 m2 plots) until reaching the known stand age of burn from the combustion database. An ignition event was then initiated at this year on the day of burn from the observations. Because UVAFME simulates daily weather by drawing from historical (1960–1990) climate distributions, an ignition event may occur directly after a simulated rainfall event, when in reality it will likely have occurred during a dry period. Thus, we additionally used the fuel moisture codes (FFMC and DMC) from the CFFDRS and wind speed for the day of burn, which were also obtained from the combustion database. These data were derived from the global fire weather database and calculated using the CFFDRS (Field et al 2015, Walker et al 2020a). Simulated aboveground and belowground combustion from these fires, averaged by ecoregion, were then compared to those observed in Walker et al (2020a). T-tests were conducted for each ecoregion grouping to determine if there were significant (p < 0.01) differences between observed and modeled values. Aboveground fuels in UVAFME were classified as surface litter (i.e. needles, leaves, and coarse woody debris), boles, and live fuels. Belowground fuels were classified as humus and live and dead roots.
2.4. Climate change analysis
Following model updates and testing, we performed a climate change application at the combustion sites within Alaska and the Northwest territories (NWT) (n = 300; table 1; figures 1 and S1, S2 in the supplementary material) to predict how forest biomass, species composition, and fire regimes (e.g. intensity, probability, and severity) may shift under changing climate. Only these sites were chosen because predictions of future lightning strike frequencies were only available for Alaska and the NWT (Veraverbeke et al 2017). Future climate data (mean monthly temperature and precipitation) were obtained from a 771 m resolution CMIP5-model average for RCP 4.5 (moderate) and 8.5 (extreme) scenarios for 2006–2100 (Leonawicz et al 2015a, 2015b).
Table 1. Average climate change forcing (years 2006 through 2100) for 300 sites within Alaska and the NWT.
| Region | Ecoregion | Scenario | Mean temperature change (°C) | Mean precipitation change (mm) | Mean lightning strike change (strikes km−2 d−1) |
|---|---|---|---|---|---|
| Alaska | Alaska Boreal Interior | RCP 4.5 | +3.27 | +67.87 (+24%) | +1.23 × 10−4 (+68%) |
| RCP 8.5 | +6.5 | +95.3 (+34%) | +2.45 × 10−4 (+135%) | ||
| Boreal Cordillera | RCP 4.5 | +2.92 | +68.82 (+19%) | +2.46 × 10−4 (+60%) | |
| RCP 8.5 | +5.85 | +113.7 (+31%) | +4.97 × 10−4 (+121%) | ||
| Northwest Territories | Taiga Plain | RCP 4.5 | +2.83 | +14.96 (+5%) | +7.92 × 10−5 (+24%) |
| RCP 8.5 | +6.19 | +92.18 (+32%) | +1.74 × 10−4 (+52%) | ||
| Taiga Shield | RCP 4.5 | +3.06 | +24.49 (+9%) | +2.10 × 10−5 (+26%) | |
| RCP 8.5 | +6.69 | +83.65 (+31%) | +4.60 × 10−5 (+57%) |
For this application, fire ignitions were based on input lightning strike frequency, and fuel moisture was dynamic and based on climate forcing data. We simulated increasing lightning strike frequency based on predictions from Veraverbeke et al (2017) (see supplementary material section 3). For these climate change simulations, we ran UVAFME at the 300 sites from bare ground initiation (each with 200 plots at 500 m2) to the inventory stand age in 2006 and subsequently initiated the climate change application. To determine the effect of climate on changing forest and fire conditions, we ran a control simulation for the same time period, sites, and number and size of plots, but continued to use historical (1960–1990) climate drivers. Model output was averaged across ecoregions (Alaska Boreal Interior and the Boreal Cordillera for sites within Alaska, and the Taiga Plain and Taiga Shield within the NWT).
3. Results
3.1. Comparison to observational data
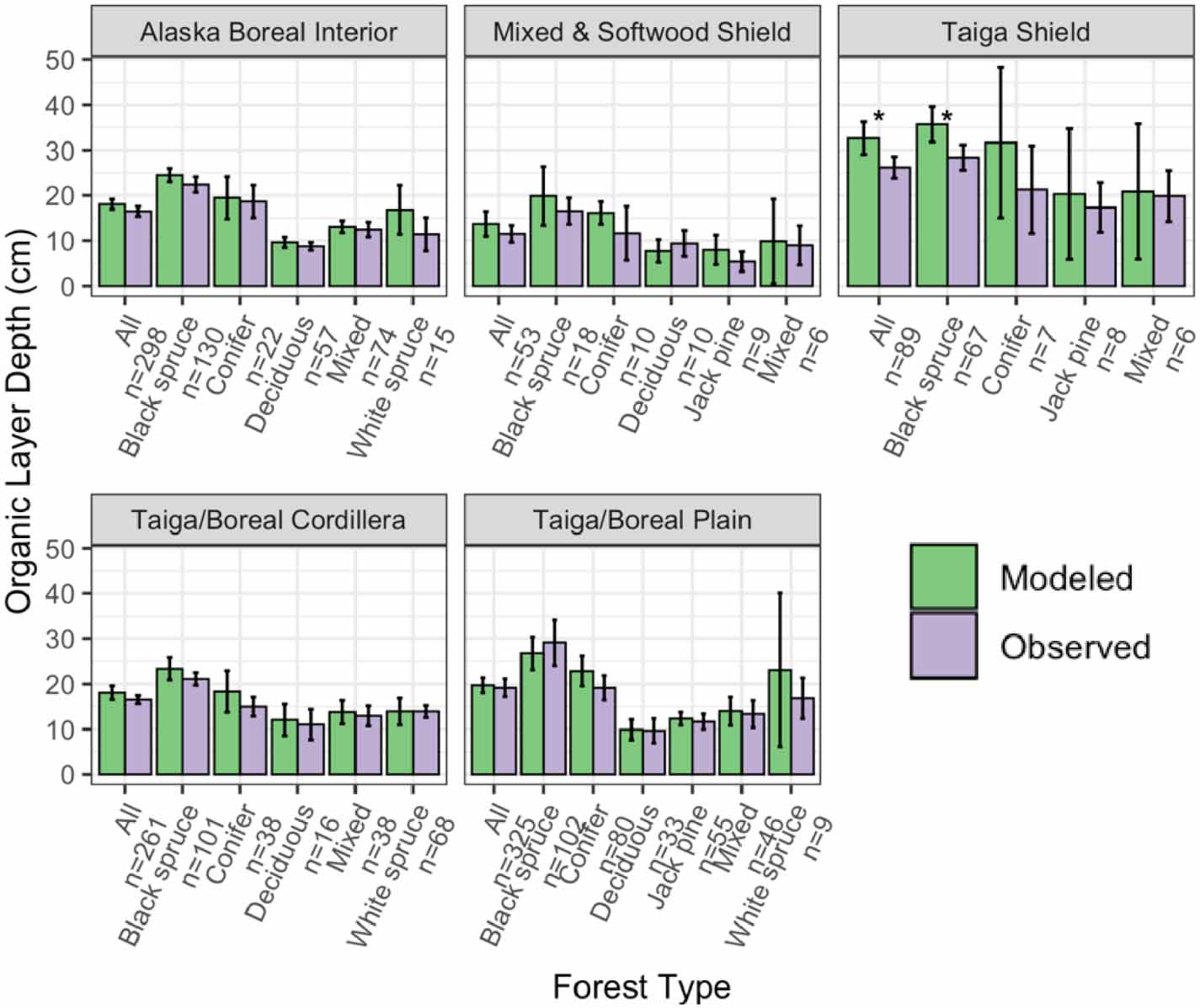
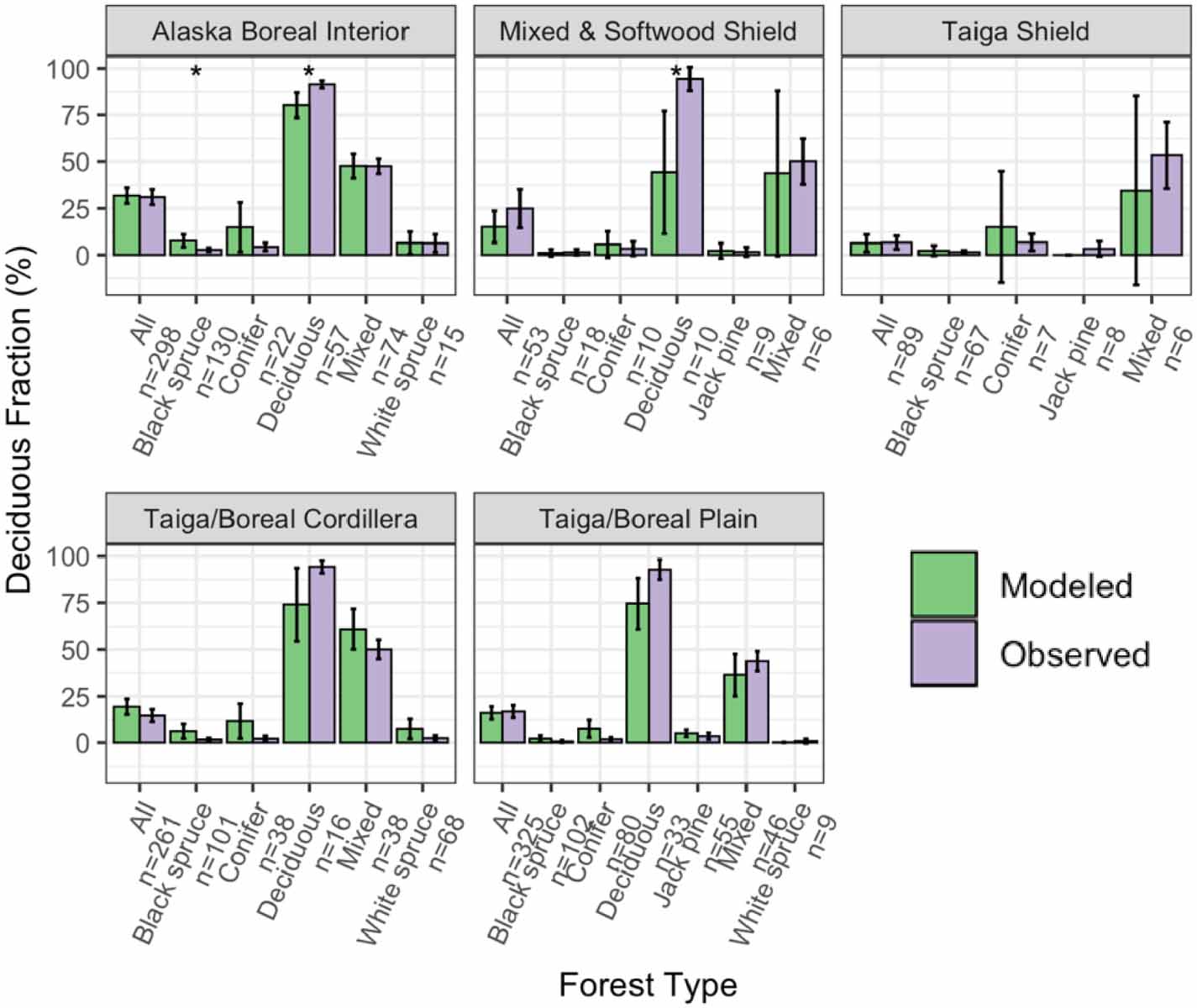
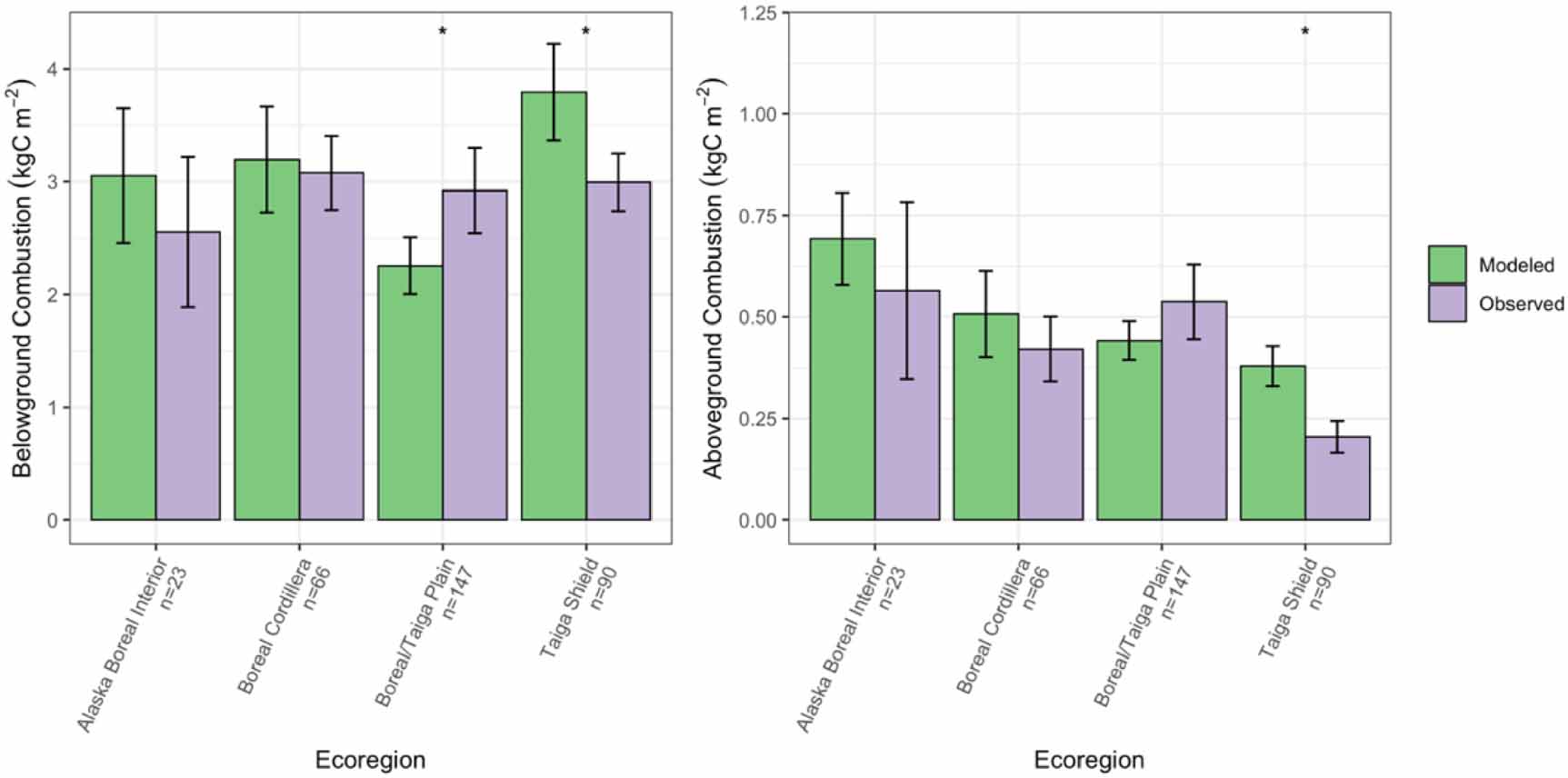
Overall, total basal area compared well between the model-simulated and observed averages across ecoregions and forest types (RMSE = 4.30 m2 ha−1; figure 2). Although in many ecoregions UVAFME underpredicted total basal area, especially that of the Mixed and Softwood Shield and the Taiga Shield, only four of the 30 groups had differences that were statistically significant (p < 0.01). Specifically, UVAFME underpredicted basal area overall ('All') and for Jack pine stands in the Taiga/Boreal Plain and overall ('All') and in black spruce stands in the Taiga Shield. In contrast, UVAFME overpredicted basal area within white spruce stands in the Taiga/Boreal Plain, although model variability was high and the sample size was low (n = 9). Model-simulated average organic layer depth also compared well with observations (RMSE = 3.61 cm; figure 3). UVAFME overpredicted organic layer depth for black spruce stands in the Taiga Shield, but overall, the model captured the organic layer depths across these forest types and ecoregions well. Simulated deciduous fraction was less well constrained, with high variability across sites (RMSE = 13.2%; figure 4). Deciduous fraction was underpredicted in deciduous stands in the Mixed and Softwood Shield and Alaska Boreal Interior, and was slightly overpredicted in the black spruce forest type in Alaska Boreal Interior. Within most other forest types, modeled deciduous fraction was not significantly different (p > 0.01) from the observed inventory data (figure 4). Simulated above- and below-ground combustion also compared well to observations (RMSE = 0.13 kgC m−2 and 0.58 kgC m−2, respectively; figure 5). There was some overprediction of combustion in the Taiga Shield and underprediction in the Boreal/Taiga Plain; however, overall, the model updates resulted in good agreement between simulated and observed combustion within the region (figure 5).
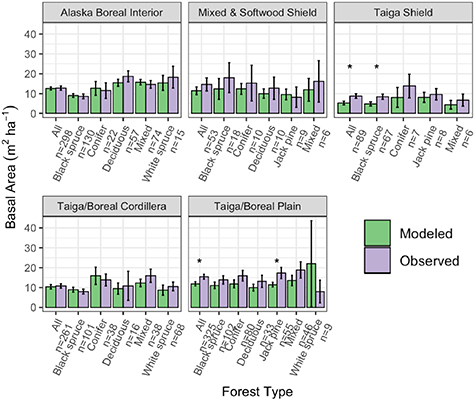
Figure 2. Comparisons of modeled and observed total basal area across inventory sites, grouped by forest type and ecoregion. Error bars indicate 95% confidence intervals. Asterisks indicate significant (p < 0.01) differences between modeled and observed values based on individual t-tests.
Download figure:
Standard image High-resolution image
Figure 3. Comparisons of modeled and observed organic layer depth across inventory sites, grouped by forest type and ecoregion. Error bars indicate 95% confidence intervals. Asterisks indicate significant (p < 0.01) differences between modeled and observed values based on individual t-tests.
Download figure:
Standard image High-resolution image
Figure 4. Comparisons of modeled and observed deciduous fraction by basal area across inventory sites, grouped by forest type and ecoregion. Error bars indicate 95% confidence intervals. Asterisks indicate significant (p < 0.01) differences between modeled and observed values based on individual t-tests.
Download figure:
Standard image High-resolution image
Figure 5. Comparisons of modeled and observed above- and below-ground combustion across the observational database, averaged by ecoregion. Error bars indicate 95% confidence intervals. Asterisks indicate significant (p < 0.01) differences between modeled and observed values based on individual t-tests.
Download figure:
Standard image High-resolution image3.2. Climate change analysis
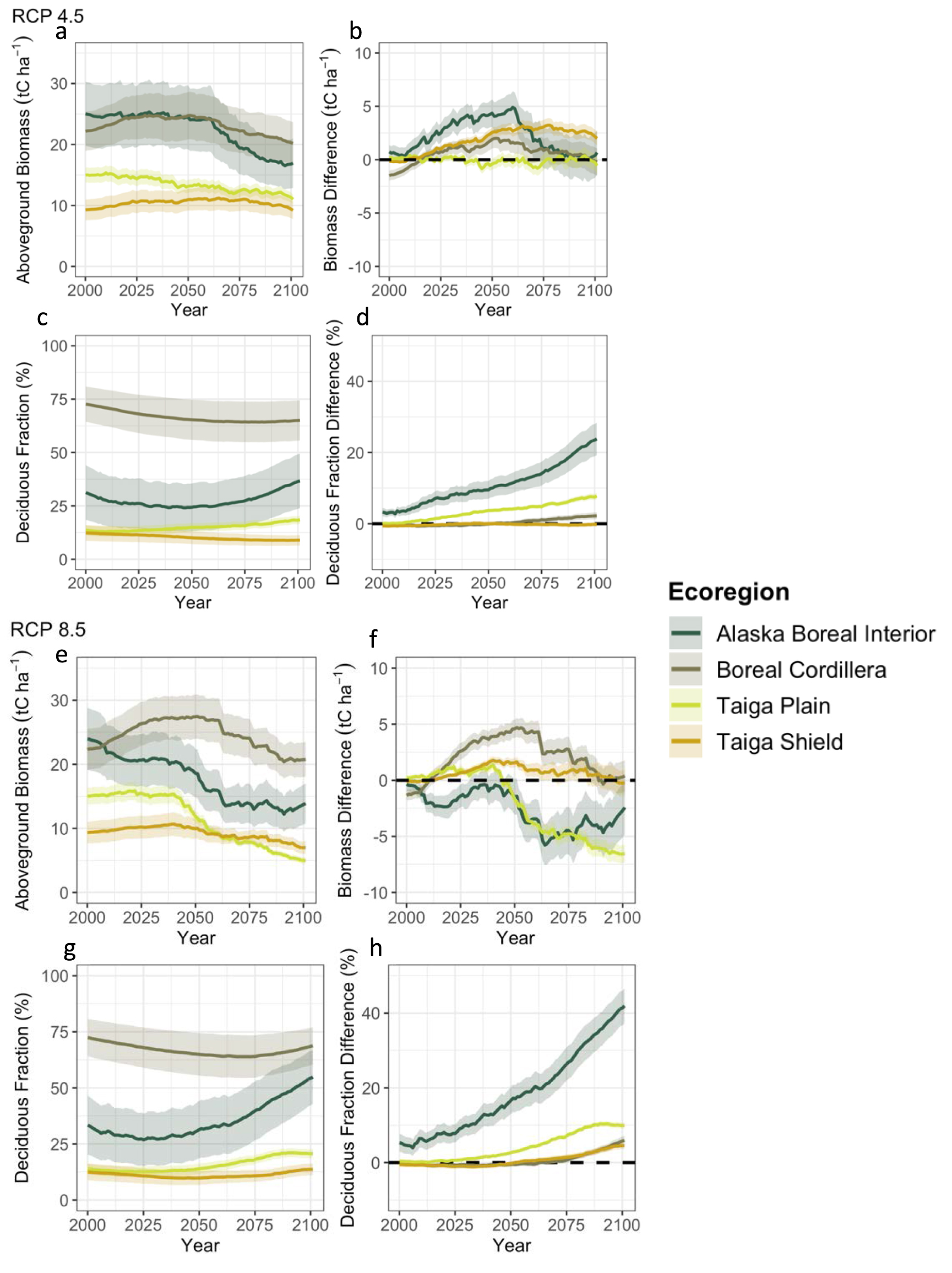
At the sites (n = 300) where climate change was applied, biomass and deciduous fraction change from 2006 to 2100 varied by ecoregion and by RCP scenario (figure 6). Under the more moderate climate change (RCP 4.5) scenario, there was relatively little change in aboveground biomass in absolute terms (figure 6(a)) and as compared to the control simulation (figure 6(b)) at any of the sites, corresponding to little change in moisture stress conditions for these sites (figure S3 in the supplementary material). Deciduous fraction initially (until ∼2050) decreased or remained relatively constant for all ecoregions (figure 6(c)). Deciduous fraction increased in Alaska Boreal Interior after about 2050, and increased gradually throughout the simulation in the Taiga Plain. Deciduous fraction was also higher in Alaska Boreal Interior and the Taiga Plain as compared to the control simulation without climate change (figure 6(d)). Under more extreme climate change (i.e. RCP 8.5), aboveground biomass decreased as a result of climate change for Alaska Boreal Interior and the Taiga Plain, but increased or remained relatively constant for the Taiga Shield and Boreal Cordillera (figures 6(e) and (f)). As in RCP 4.5, deciduous fraction initially decreased or remained relatively constant for the first 25–75 years of the RCP 8.5 climate change application, depending on the ecoregion (figure 6(g)). By 2100, all ecoregions had higher deciduous fraction as compared to the control simulation without climate change (figure 6(h)). Increases in deciduous fraction occurred more quickly and were greatest for Alaska Boreal Interior. Organic layer depth decreased for all but the sites within the Taiga Shield under RCP 8.5, and thaw depth increased for all ecoregions under both RCP 4.5 and RCP 8.5 (figure S4).

Figure 6. Model-simulated average aboveground biomass and deciduous fraction by biomass (left) as well as difference from control simulation with historical climate (right) for an RCP 4.5 scenario (top) and RCP 8.5 scenario (bottom) for 300 sites within Alaska and the Northwest Territories, averaged by ecoregion. Shaded regions show 95% confidence intervals.
Download figure:
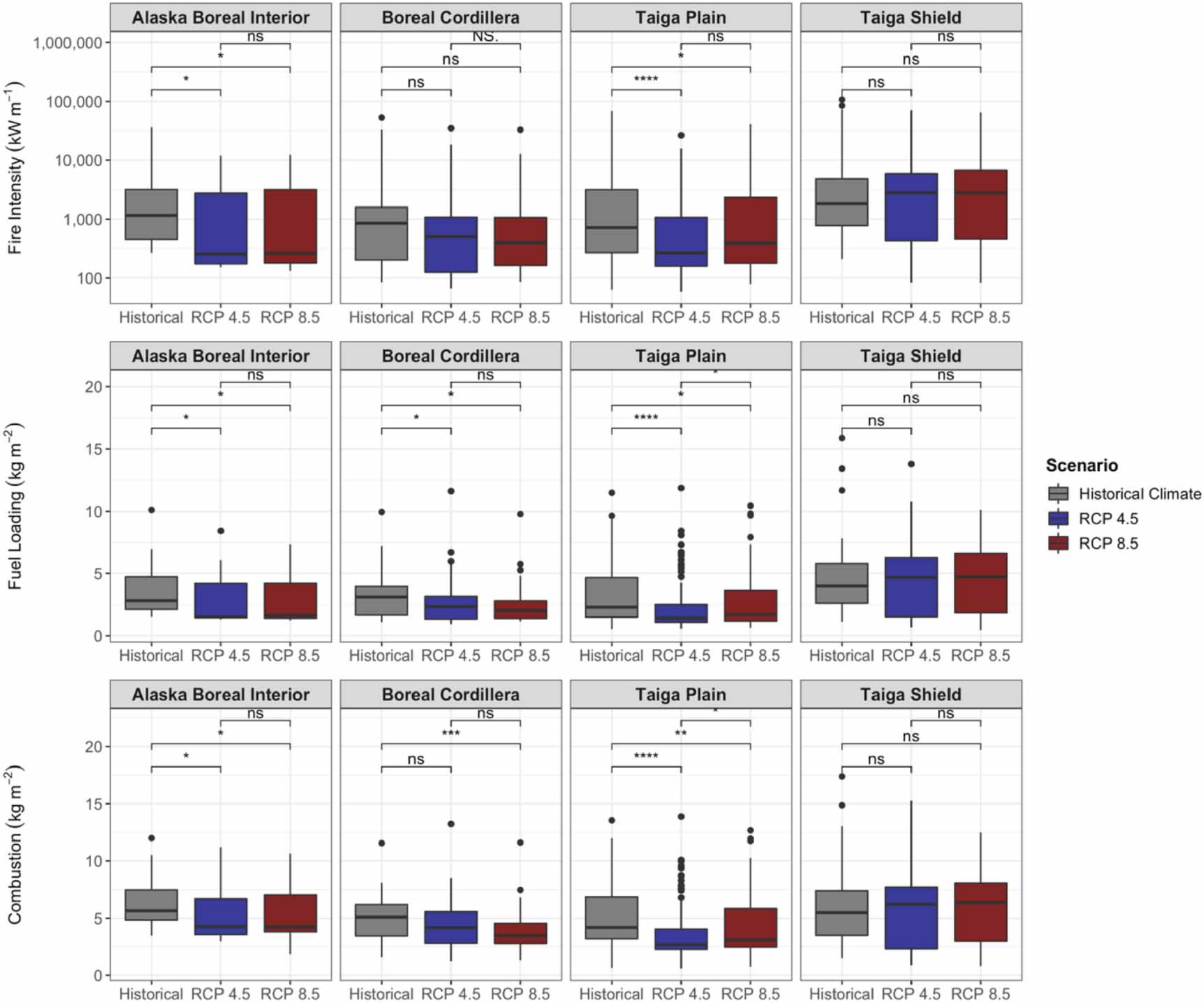
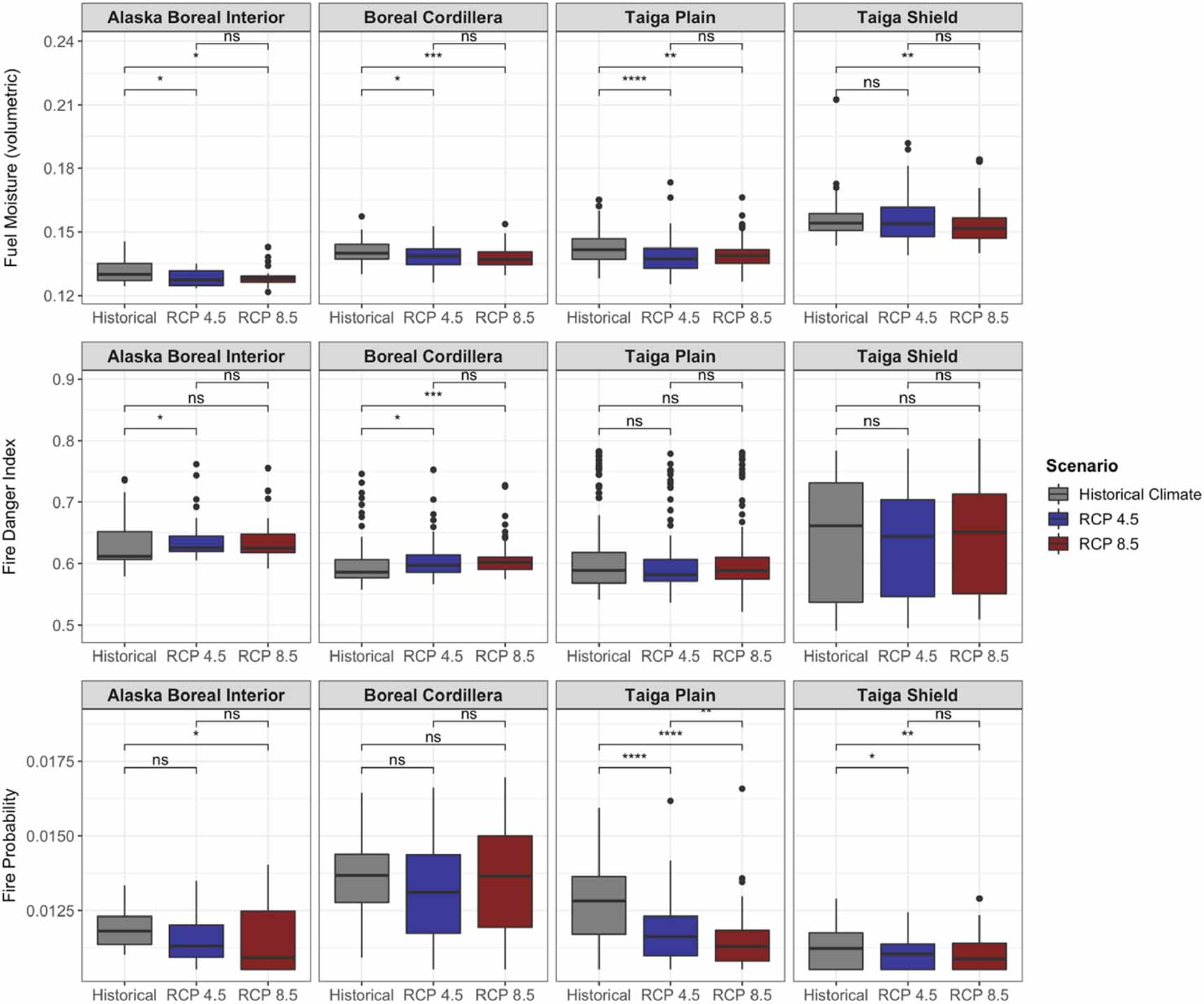
Standard image High-resolution imageDifferences in fire severity, intensity, and probability from the control simulation with historical (1960–1990) climate also varied by ecoregion and climate change scenario (figures 7 and 8). Fire intensity and fire severity (i.e. total combustion) on average decreased with climate change for Alaska Boreal Interior, the Boreal Cordillera, and the Taiga Plain, although this decrease was not statistically significant for most comparisons in the Boreal Cordillera (figure 7). In sites where deciduous fraction increased by at least 30%, the decline in fire intensity with climate change was large and statistically significant, whereas in sites where there was relatively little change in deciduous fraction (<5%), fire intensity did not change (figure 9). Total fuel loading (i.e. needles/leaves, twigs, branches, moss, and small shrubs/trees) also decreased with climate change for all ecoregions except the Taiga Shield (figure 7). In the Taiga Shield, simulated fire intensity, combustion, and fuel loading increased with climate change, although these differences were not statistically significant. Most comparisons showed non-significant differences between the RCP 4.5 and RCP 8.5 fire regimes. Notably, although average intensity and combustion decreased for most ecoregions as a result of climate change, maximum values remained consistent, indicating the possibility for extreme fire events in the future. Fuel moisture decreased for all ecoregions for both RCP 4.5 and RCP 8.5 (figure 8), although the FDI (equation (S25) in the supplementary material) increased for some of the ecoregions—Alaska Boreal Interior and the Boreal Cordillera in particular (figure 8). Although fuel drying persisted as a result of climate change, fire probability (calculated as the number of fires occurring at each site during the simulation divided by the number of years in the simulation) decreased for most ecoregions (figure 8).

Figure 7. Model-simulated distributions of fire intensity (kW m−1), fuel loading (kg m−2), and combustion (kg m−2) for 300 sites across different ecoregions in Alaska and NWT for historical, RCP 4.5, and RCP 8.5 climate scenarios for years 2006 through 2100. Asterisks denote significance level of Kruskal–Wallis tests (ns: p > 0.05; *: p ⩽ 0.05; **: p ⩽ 0.01; ***: p ⩽ 0.001; ****: p ⩽ 0.0001). Note the log-10 scale for fire intensity.
Download figure:
Standard image High-resolution image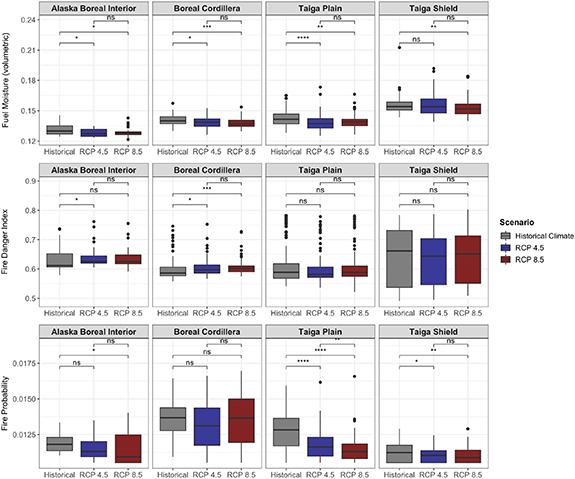
Figure 8. Model-simulated distributions of fuel moisture (volumetric), FDI (0–1) and fire probability (0–1) for 300 sites across different ecoregions in Alaska and NWT for historical, RCP 4.5, and RCP 8.5 climate scenarios for years 2006 through 2100. Asterisks denote significance level of Kruskal–Wallis tests (ns: p > 0.05; *: p ⩽ 0.05; **: p ⩽ 0.01; ***: p ⩽ 0.001; ****: p ⩽ 0.0001).
Download figure:
Standard image High-resolution image
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 9. Model-simulated distributions of fire intensity (kW m−1) for simulated sites that had a greater than 50% increase in deciduous fraction (right) and for sites that had little deciduous fraction change (left) for historical, RCP 4.5, and RCP 8.5 climate scenarios for years 2006 through 2100. Asterisks denote significance level of Kruskal–Wallis tests (ns: p > 0.05; *: p ⩽ 0.05; **: p ⩽ 0.01; ***: p ⩽ 0.001; ****: p ⩽ 0.0001). Note the log-10 scale on the y-axis.
Download figure:
Standard image High-resolution image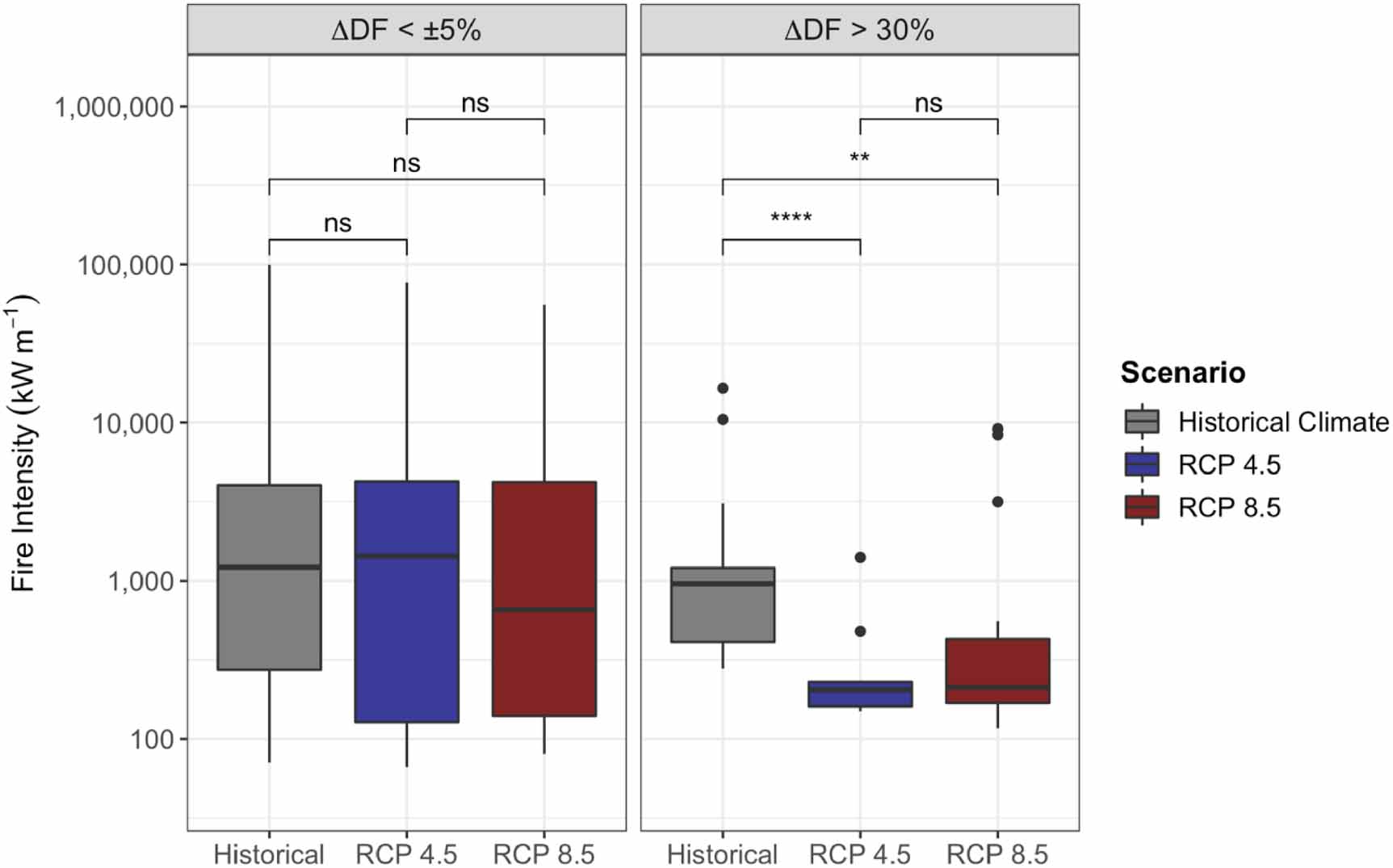{kind=link}
4. Discussion
Following on validation efforts from Foster et al (2019) within Interior Alaska, the tests presented here across Alaska and western Canada confirm UVAFME's ability to predict forest conditions, organic layer depth, and combustion within western boreal North America (figures 2–5). Although there were some discrepancies between modeled and observed basal area, organic layer depth, and deciduous fraction in some forest types, in general the model performed well at predicting forest conditions and drivers across a broad range of site types. Discrepancies in deciduous fraction (figure 4) could have occurred because only sites with an observed stand age less than 100 years had species available for recruitment based on inventory data; otherwise, all species present in species range maps for that location were allowed to establish and grow at older sites (see supplementary material section 3.3). Additionally, initial (i.e. bare-ground) species-specific seed density was not initialized with any disturbance or other site history in mind. In the North American boreal forest, local-scale species composition is highly dependent on fire history (Chapin III et al 2006a). Because we did not initialize seed banks with consideration for each site's fire history or current species composition, it is possible that the initial modeled seed and seedling banks were skewed away from their realistic values. Serotinous cones and the effect of fire on seed rain and seedling regeneration are present in UVAFME (Foster et al 2019), but we do not currently consider these effects at stand initiation. In the future, initialization of species-specific seed and seedling banks will be included in UVAFME to better understand how these processes impact stand-level forest dynamics.
The climate change results at the 300 sites (figures 6–9) indicate the importance of dynamic vegetation and fuels when predicting future forest and wildfire conditions. Simulated vegetation response to shifting climate differed based on ecoregion (figure 6) and physiographic condition (figures S5 and S6), with drier sites showing overall decreases in biomass due to drought stress (figure S7), and wetter sites showing overall increases in biomass with climate change, as in Foster et al (2019). These changes in vegetation are also similar to what has been predicted in other vegetation modeling studies in boreal North America (Mekonnen et al 2019, Hansen et al 2021). A similar pattern was predicted with changing fire intensity across different physiographic conditions (figure S8): drier sites, which also tended to have a higher deciduous fraction and greater decreases in biomass, had lower fire intensity with climate change than wetter sites. At these dry sites, increasing temperatures and drying conditions (figure S3) caused decreases in biomass and/or shifts in species composition to more drought-tolerant species such as quaking aspen (Populus tremuloides) (figures 6 and S6), leading to less flammable fuels (Johnson 1992, Krawchuk et al 2006). In addition, decreases in organic layer depth and surface fuel loading (figures 7 and S4) lowered the overall intensity and severity of fires at many of the sites as climate change progressed. This decline in fire intensity was mostly linear over time (figure S9), and was strongest at sites where deciduous fraction increase was greatest (figure 9), indicating that while fuel drying may be important, and did occur at these sites (figure 8), the bottom-up, negative feedback on fuel loading and geometry (i.e. bulk density and SAV) outweighed the increasing flammability from drying fuels and increased lightning strike frequency.
The more extreme climate change scenario (RCP 8.5) also resulted in a decline in Jack pine (Pinus banksiana) biomass within the Northwest Territories (the Taiga Plain and Taiga Shield ecoregions) (figure S10), likely due to an increase in moisture stress (figure S7), even though Jack pine is classified as drought-tolerant (Burns and Honkala 1990). Despite little change in deciduous fraction, fuel loading and ultimately fire intensity also declined across these sites (figure S11), driven by declining Jack pine biomass and rapidly decaying fuels. However, fire probability increased across these sites, indicating a shift towards smaller, more frequent fires. In contrast, UVAFME predicted little change in deciduous fraction and even increased biomass in the Taiga Shield compared to the control simulation (figure 6), accompanied by increasing fire intensity and severity (figure 7), although these differences were not significant. These differential responses of vegetation and wildfire regimes based on ecoregion, physiographic condition, and deciduous fraction trajectory highlight the importance of simulating both dynamic fuels and dynamic vegetation, and their interactions, when predicting future wildfire regimes. In general, UVAFME predicted stronger bottom-up controls and negative feedbacks on wildfire severity and intensity under climate change, ultimately driven by these key linkages between vegetation, fuels, and fire.
The projected precipitation changes impacted both the simulated vegetation and wildfire response and drove differences in the vegetation response to climate change across ecoregions. In the Boreal Cordillera, annual precipitation was highest and increased under the RCP 8.5 climate scenario (figure S2, table 1), resulting in little change in moisture conditions or deciduous fraction (figures S3 and 6). Changes in precipitation are difficult to predict, especially at the global climate model scale (Soja et al 2007, Pendergrass et al 2017), and these uncertainties can propagate to simulated vegetation dynamics (Bonan et al 2019). The simulations presented here additionally did not include the potential effect of CO2 fertilization on tree growth, which may have mitigated some mortality and declining productivity due to drought stress had it been included (Sullivan et al 2017). However, it is currently unclear the extent to which CO2 fertilization will mitigate future climate stressors (Norby et al 2015). Thus, our results should be considered alongside uncertainties in climate change forcing data, as well as the potential mitigating effect of CO2 fertilization.
Our findings are similar to what has been found in other fire self-regulation studies in the boreal region (Johnstone et al 2011, Krawchuk and Cumming 2011, Girardin et al 2013, Hart et al 2019, Walker et al 2020b), with some key differences. Krawchuk and Cumming (2011) and Hart et al (2019) found negative impacts of stand composition and forest structure on fire initiation, burned area, and fire probability; however, in these studies, negative feedbacks on fuel were not sufficient to reduce climate-induced increases in future fire activity. Johnstone et al (2011) used the landscape model ALFRESCO to investigate the effect of fire severity and climate change on vegetation-wildfire feedbacks and found a negative feedback of increasing deciduous cover on future fire hazard following severe fires. However, these negative feedbacks were not strong enough to compensate for increased fire activity overall due to climate warming and fuel drying (Johnstone et al 2011).
The simulations with ALFRESCO and those in Hart et al (2019) and Krawchuk and Cumming (2011) did not include dynamic fuels or the possibility for sustained deciduous dominance under climate change, whereas ours with UVAFME do. Thus, our results highlight the importance of bottom-up feedbacks between vegetation, fuel conditions, and wildfire activity. It is also important to note that ALFRESCO is a landscape model capable of simulating fire spread across simulated grid cells, whereas this version of UVAFME does not the include spatial propagation of fire. Much of the negative feedback simulated by ALFRESCO arose from fewer contiguous patches of spruce and thus interruptions of fire spread by less flammable deciduous patches (Johnstone et al 2011). This landscape-scale propagation of fire and potential increase in burned area with climate change are important components of wildfire dynamics in the boreal zone (Flannigan et al 2009, Krawchuk et al 2009). Although our results with UVAFME show the importance of bottom-up controls on fire initiation, intensity, and severity at the site scale, top-down feedbacks of increased lightning strikes, fuel drying, and longer fire seasons on fire propagation and landscape heterogeneity will also impact future wildfire regimes. Going forward, models that incorporate dynamic fuels and vegetation (e.g. UVAFME), as well as the spatial propagation of fire and spatial impacts of fire on seed source availability (e.g. iLand, Hansen et al 2021) would best be used, separately or in concert, to determine where and the extent to which these bottom-up and top-down controls will be strongest.
5. Conclusions
Wildfire regimes in the North American boreal forest are the consequence of feedbacks between biological vegetation and fuel dynamics, as well as geophysical feedbacks between climate, fire, and the carbon cycle. Our results show that bottom-up controls on vegetation composition and fuel type and loading are as or more important than top-down controls of climate warming within the boreal forest. We found evidence for declining biomass within interior Alaska and the Taiga Plain of the Northwest Territories, as well as increasing deciduous fraction as a result of increasing temperatures. Where deciduous fraction increases and biomass and fuel declines were greatest, we found strong evidence for decreasing fire intensity and severity despite increased fuel drying.
Dynamic vegetation and dynamic fuels are crucial for accurately capturing the competing differences between top-down and bottom-up controls on future fire regimes. Boreal forests are a major component of the global carbon and energy cycles, and changing boreal fire regimes will impact regional as well as global climate through changing terrestrial carbon stores, albedo, and emissions. Nonetheless, Wang et al (2021) found that Earth system models in the CMIP6 ensemble overpredicted boreal forest aboveground biomass in comparison to that derived from satellite observations, in part due to missing or inadequate representations of fire disturbance. Moreover, Archibald et al (2018) cite dynamic and accurate fuels as well as biological vs. climate drivers as two key research challenges to modeling fire within the Earth system. Our study emphasizes the importance of these processes and provides methods and impetus for their inclusion in large-scale dynamic vegetation models.
Acknowledgments
This work was supported by NASA Arctic Boreal Vulnerability Experiment (ABoVE) Grant 80NSSC19M0112 and DoD SERDP contract RC18-1183. Model simulations were run on Northern Arizona University's Monsoon computing cluster, funded by Arizona's Technology and Research Initiative Fund. We thank Environment and Climate Change Canada for their generous permission to use Canadian Lightning Detection Network data.
Data availability statement
The data that support the findings of this study are openly available at the following URL/DOI: https://github.com/UVAFME/UVAFME_model.