

Abstract
Pure and even-aged (PEA) forests of fast-growing conifer species have for a long time been key providers of industrial raw material. Despite recent concerns regarding their greater sensitivity to major natural disturbances, their impacts on biodiversity and their funding efficiency, PEA conifer forests could remain a major economic target given the ongoing European strategy for bioeconomy. These forests are encountered in contrasted climates and in both native and introduced contexts across Europe, giving rise to high uncertainties regarding their growth responses to current climate change. Using the French National Forest Inventory data, we studied the radial growth of eight major conifer species in European forestry across 16 regional native and introduced PEA forest systems (n > 10 000 trees). Growth trends over the 2006–2016 period exhibited a significant negative association with the absolute growth level in 2006, with the strongest negative growth trends found for emblematic PEA forestry species (e.g. Norway spruce and Douglas fir), and the strongest positive trends for pine species (e.g. Scots pine). While the greater growth rate advantage of some species may shorten rotation and lower risk exposures for future decades, their recent lowered productivity may affect the forest sector in the long run. The prevalence of PEA forests across European forest landscapes and their increasingly reported lower resilience to climate change compared to more complex forest systems call for the establishment of a long-term European forest policy strategy. Maintaining the environmental, social and economic benefits of forests should remain a priority in the European agenda, regardless of the financial costs at stake.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. The era of fast-growing conifer forests
Over the past two centuries, fast-growing conifer species, under the silvicultural paradigm of intensive pure and even-aged (PEA) forestry [1], have become a key provider of raw material for the timber industry owing to their productivity, regular morphology and wood anatomy. To further increase productivity potential and bypass native growth constraints, the forest sector has not only introduced conifer species outside their natural range but also exotic species [2]. Afforestation programs have mainly transferred cold-adapted species into warmer regions and drought-adapted species into cooler/wetter regions. The interest in fast-growing conifer species has also pushed breeding programs towards a continuous genetic improvement of productivity rates, resistance to pathogens and material quality (e.g. [3, 4]). Pro-active funding policies have further supported the establishment of monocultures of fast-growing conifers, increasing their prevalence across European forest landscapes [5, 6]. Despite recent concerns regarding a higher sensitivity to major natural disturbances [7], impacts on biodiversity [8] and funding efficiency [2], PEA conifer forestry [9] could remain a major forest type given the ongoing European strategy for bioeconomy [10]. Whether climate change constitutes a detrimental threat to PEA forestry and more largely to the European forest sector now comes into debate (e.g. [11, 12]).
2. Conifers in France, a demonstrative case study
In France, PEA conifer forests are encountered across contrasting climatic contexts (oceanic, continental and Mediterranean; figure 1(A)) and account for 25% and 40% of the forest area and growing stock, respectively. Over the past 150 years, afforestation programs have triggered a doubling of their spatial extent. Norway spruce (Picea abies Karst.) and Scots pine (Pinus sylvestris L.), two cold-adapted species native to the French Alps, have been largely introduced at low and mid-elevations (e.g. Northern Plains and Massif Central). Maritime pine (Pinus pinaster Ait.), native to northern Africa, has also been intensively introduced; firstly in the South West, where it has turned into a French emblem of large-scale intensive forestry known as 'Forêt des Landes', and then across the Northern Plains. Corsican pine (Pinus nigra subsp. laricio Maire), a drought-adapted pine species native to the Mediterranean region, has been introduced in the cooler and wetter Northern Plains. Finally, Douglas fir (Pseudotsuga menziesii Mirb. Franco), native to North America, has more recently been massively introduced at low and mid-elevations (Northern Plains and Massif Central), to the extent that France presents the second highest growing stock of Douglas fir after the USA. These features make this western European context a perfect case study for observing how PEA conifer forests in native and introduced contexts react to ongoing climate change.

Figure 1. Recent growth trendsof PEA conifer forests in France under climate change. (A) Geographical location and relief characteristics of bioclimatic regions under study. (B) Association between radial growth trends over the 2006–2016 period and average radial growth in 2006 across the 16 regional PEA forest systems in focus. Regional forest systems are labeled using their corresponding regional acronym in panel (A). Native and introduced forest systems are displayed using circles and squares, respectively. Standard deviations (95%) of native and introduced contexts along the x and y spaces are plotted using continuous and dashed segments, respectively. The black line illustrates the linear regression between regional growth trends and average regional radial growth with all forest systems included. The R2 and P value of this regression are displayed in the figure. Colored lines represent the same linear regression but at intra-species level when possible (more than one regional forest system studied for a given tree species). Radial growth anomaly chronologies from which trends were computed are presented in figure S1.
Download figure:
Standard image High-resolution image3. Isolating climate effects on growth using National Forest Inventory data
Using the French National Forest Inventory sampling design and data, we studied the radial growth of eight conifer species prevalent in European forestry in PEA forests across different bioclimatic regions (n > 10 000 trees; figure 1). In total, 16 PEA regional forest systems were studied (figure 1(B)). Regional forest systems were classified as either native or introduced based on distribution maps elaborated by the European Forest Genetic Resources program (www.euforgen.org) and available historical knowledge of species introduction in new regional contexts (table S1, available online at stacks.iop.org/ERL/16/024030/mmedia). The present study completes previous research [13] by investigating further the origins of growth trend variability observed across PEA conifer systems over the 2006–2016 period through the link between growth trends and the initial growth level in 2006.
Radial increment series were filtered out from non-climatic signals at the system level using linear modeling and an ensemble of 42 predictive tree, stand, soil and long-term climate variables [13]. Remaining inter-annual radial growth variability, supposed to be primarily associated with short-term climate variability, was converted into relative growth anomalies by expressing the growth deviation of a specific year as a ratio of its residual growth level to the residual growth during the reference year 2006. Absolute radial growth chronologies were finally obtained by multiplying relative growth anomalies by the average growth level during the reference year (figure S1). The strength of the herein-developed modeling approach is that changing the reference year would neither impact the relative position of growth anomalies, nor the relative growth difference (deviation %) between any two years, nor the absolute radial growth chronology obtained (in mm) (figure S2).
Regional trends in inter-annual growth anomalies were subsequently regressed against regionally aggregated biotic and short-term seasonal climate predictors. Predictors included e.g. the Gini index, a plot-level measure of tree-size heterogeneity derived from tree diameter field measurements which value ranges between 0 (perfectly homogeneous stands) and 1 (singularly heterogeneous stands), water field capacity directly measured in the field, and averages and trends in seasonal climate computed over the 2006–2016 period using the European E-OBS 0.5° × 0.5° gridded dataset (www.ecad.eu).
4. Greatest growth declines in fast-growing systems
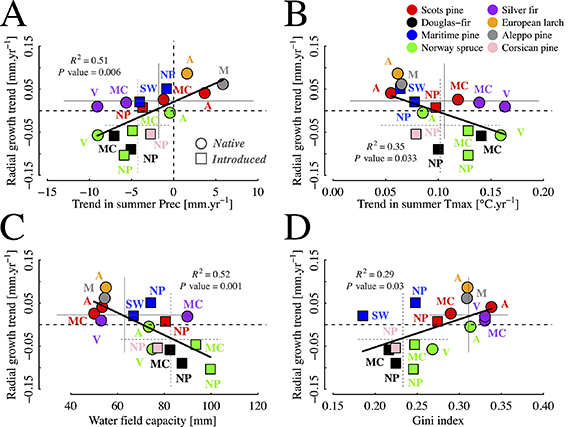
Analyses revealed that growth trends over the 2006–2016 period exhibited a negative association with the absolute growth level in 2006 (R2 = 0.61, P < 0.001; figure 1(B)), with the strongest negative trends undergone by emblematic species of the most recent afforestation programs (e.g. Norway spruce and Douglas fir, which represent the highest and second highest conifer growing stock in France, respectively; www.inventaire-forestier.ign.fr). As a major finding, this pattern was also observed across species and across regional samples within species (figure 1(B)) suggesting that climate-driven transient conditions of tree growth affect tree-specific populations in a universal direction (e.g. regression-to-the-mean process [14]), with fast-growing forest systems showing systematically more detrimental growing trajectories than slow-growing systems regardless of the biological organization level in focus. Causal explorations, presented in the previous study [13], indeed revealed that negative growth trends were significantly accentuated by decreasing summer precipitation (R2 = 0.51, P = 0.006; figure 2(A)) and increasing summer temperature (R2 = 0.35, P = 0.033; figure 2(B)), giving ground to adverse climate change effects on PEA conifer forests, especially in introduced contexts. Yet, consistent with the exhibited trade-off (figure 1(B)), greater growth declines were found in forest systems with greater water field capacity (R2 = 0.52, P = 0.001; figure 2(C)), providing a resource-oriented ground to greater growth declines at greater initial growth. Lastly, forest systems with greater stand diameter heterogeneity exhibited greater growth (R2 = 0.29, P = 0.03; figure 2(D)).
{kind=link}
Figure 2. Major environmental indicators of the 2006–2016 growth trends of the 16 regional PEA conifer forest systems under study. Native and introduced forest systems are displayed using circles and squares, respectively. Regional forest systems are labeled using their corresponding regional acronym in figure 1(A). Standard deviations (95%) of native and introduced systems along the x and y spaces are plotted using continuous and dashed segments, respectively. Trend in summer precipitation (A) and maximum temperature (B) were computed over the 2006–2016 period. The black line illustrates the overall linear regression between growth trends and the regionally aggregated predictive variable in focus. The R2 and P value of this regression are displayed in each panel. The results presented in the different panels are reprinted from [13], Copyright (2020), with permission from Elsevier.
Download figure:
Standard image High-resolution image{kind=link}
5. Climate–forest interactions: on the need to rethink silvicultural paradigms
Over a period of generalized increasing thermal (between +0.55 °C and +1.72 °C in 11 years depending on PEA systems; figure 2(B)) and water constraints (figure 2(A)), greater climatic stress to tree growth is hence being observed in introduced systems where growth (figure 1(B)) is not limited by soil water potential (figure 2(C)) but where competition for resources is greater (figure 2(D)). The ongoing and forecasted pace of climate change will exacerbate climatic constraints, thereby contracting the optimal geographical envelopes of many conifer tree species [15]. Even though greater initial growth potential may persist for a few decades (figure 1(B)) and facilitate the shortening of rotations and reduction of forest exposure to sudden disturbance and stress, it may also yield a gradual decrease in productivity (figure 1(B)) with potential major impacts on the forest sector in the long term. The ongoing and forecasted impacts of climate change onto forest growth hence praise for a shift in the dominant silvicultural paradigm.
The current forestry interests in fast-growing tree species and homogeneous forest structure should be urgently revised. Renewed attention on long-term conifer species selection is needed, with slow-growing (e.g. hardy pine) species as possible primary targets due to their greater resilience to climatic stress (figure 1). Although promoting stand heterogenization may appear as a path forward to enhance the climatic resilience of PEA forest systems [16], arising technical difficulties (e.g. organizing plantation waves and selective harvesting), costs and time horizons of implementation make this solution very unlikely to happen on a large geographical scale. Furthermore, the recently highlighted greater resilience of more complex forest ecosystems [17] rather argues for an urgent amplification of initiated efforts to convert PEA forests towards such systems [11]. Adaptive forest management as a way to optimize resource use and tree-to-tree interactions may provide guiding management principles [18, 19]. We are aware that the implementation of these new paradigms requires substantial effort. Nevertheless, recent climate projections open up the possibility of a 5 °C warming by the end of the century [20] and seriously bring into question the future of European forests and of the environmental, social and economic benefits they provide. The role of a European political forest strategy is here heavily stressed to build a climate-resilient forestry (see [12]) and maintain forest services.
Acknowledgments
This study was financed by the French National Research Agency (ANR) through the 'Laboratoire d'Excellence ARBRE' and the PIA 'Lorraine Université d'Excellence' projects (ANR-11-LABX-0002-01 and ANR-15-IDEX-04-LUE, respectively) and by the Région Grand Est. We are thankful to the two anonymous reviewers who provided thoughtful comments on previous versions of this manuscript.
Data availability statement
The data that support the findings of this study are available upon reasonable request from the authors.