

Abstract
In the northern high latitudes, alternative hypotheses with regards to how warming-related shifts in seasonality influence ecosystem productivity exist. Increased plant growth associated with a longer growing season may enhance ecosystem productivity, but shifts to earlier springs may also negatively influence soil moisture status and productivity during the peak of the growing season. Here, we analyzed nearly three decades (1982–2008) of observational records and derived products, including satellite microwave and optical imagery as well as upscaled ecosystem flux observations, to better understand how shifts in seasonality impact hydrology and productivity in the North American boreal forests. We identified a dominant adverse influence of earlier springs on peak summer forest greenness, actual evapotranspiration and productivity at interannual time scales across the drier western and central sections of the North American boreal forests. In the vast regions where this spring onset mechanism operates, ecosystem productivity gains from earlier springs during the early portion of the growing season are effectively cancelled through corresponding losses in the later portion. Our results also indicate that recent decadal shifts towards earlier springs and associated drying in the midst of the growing season over western North American boreal forests may have contributed to the reported declines in summer productivity and increases in tree mortality and fire activity. With projections of accelerated northern high-latitude warming and associated shifts to earlier springs, persistent soil moisture deficits in peak summer may be an effective mechanism for regional-scale boreal forest dieback through their strong influence on productivity, tree mortality and disturbance dynamics.
Export citation and abstract BibTeX RIS

Content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Corrections were made to this article on 23 May 2013. The PDF was replaced to correct conversion artefacts in figures 2, 3 and 4.
1. Introduction
Since the early 1970s, terrestrial ecosystems in the northern high latitudes have experienced rates of warming (∼0.3–1.0 ° C/decade) that are higher than anywhere else on earth due to strong physical feedbacks associated with changes in arctic sea-ice and snow cover (Solomon et al 2007). As a result, the growing season has progressively lengthened due to both earlier springs and delayed autumns with consequences for important ecosystem processes (Richardson et al 2013). The vast circumpolar boreal forests play an important role in the climate system (Chapin et al 2000) and the global carbon cycle (Pan et al 2011) and are thought to be particularly vulnerable to climate change, especially if important disturbance regimes (e.g. fire) cross critical thresholds (Gitay et al 2001). Early studies linking trends in satellite plant growth and atmospheric CO2 suggested increased boreal forest productivity under warming and associated lengthening of the growing season (Myneni et al 1997). However, more recent studies showed declining satellite productivity trends across large portions of the boreal forests since the early 1990s that appear to be linked to warmer and drier conditions (Angert et al 2005, Goetz et al 2005, Beck and Goetz 2011). Several field-based studies also reported fundamental changes on how boreal forests have responded to recent warming, suggesting that this biome is becoming increasingly vulnerable to warming-related shifts in important disturbance regimes including temperature-induced drought stress (Barber et al 2000), fire (Kasischke and Turetsky 2006) as well as insect outbreaks (Kurz et al 2008).
Increasing rates of drought and heat-induced tree mortality have recently emerged as global-scale phenomena (Allen et al 2010), and new reports for North American (NA) boreal forests also documented spatially extensive pattern of drought-related productivity declines (Ma et al 2012) and increases in tree mortality (Peng et al 2011) in recent decades. In this context, several drought mechanisms have been proposed for the boreal forests including warming-related increases in evapotranspiration not compensated for by increases in precipitation (Barber et al 2000, Girardin and Mudelsee 2008) and changes in atmospheric circulation pattern (blocking high-pressure systems) that favor surface drying (Skinner et al 1999). Shifts to earlier springs across northern snow-dominated ecosystems may also lead to more frequent summer droughts with potentially severe impacts on the climatologically relatively dry boreal forests. A lengthening of the evaporative period and increased snow sublimation may lead to a build up of soil water deficit over the early and middle portion of the growing season (Barnett et al 2005, Jepsen et al 2012). In addition, temporal shifts to earlier snowmelt also increase soil moisture in the early part of the season when evaporative demand is still low. Since some of this water will be lost as runoff, soil moisture is reduced in the later portion of the growing season when evaporative demand is large (Barnett et al 2005).
Two recent boreal satellite analyses (Grippa et al 2005, Kim et al 2012) provided first large-scale evidence that years with early spring arrival are indeed associated with anomalously low vegetation growth during summer consistent with hypothesized effects of water stress. Further, several local ecosystem flux studies in western NA subalpine forests showed also reductions in summer plant productivity in years with early springs and shallow snow packs (Sacks et al 2007, Trujillo et al 2012) consistent with the observation that these forests rely heavily on water from snow melt even at a significantly later stage in the growing season (Hu et al 2010).
In this study, we focus on the NA boreal forests and explore the extent at which shifts in spring onset as a mechanism of summer drought may have contributed to the recent observed declines in productivity and increases in tree mortality and fire activity. We investigate this through the combined analysis of nearly three decades (1982–2008) of climate, satellite-based vegetation greenness (expressed through the normalized difference vegetation index: NDVI) and microwave data that capture the freeze–thaw (FT) status of the surface. In addition, we analyze upscaled ecosystem flux observations of water and carbon over the same study period to further examine how variations in spring arrival influence surface hydrology and ecosystem dynamics at both interannual and decadal time scales.
2. Data sets and methods
For satellite vegetation greenness data, we used the bimonthly maximum composite NDVI version G (NDVIg) data set produced by the Global Inventory Modeling and Mapping Studies (GIMMS) group (Tucker et al 2005). The NDVI is computed as the difference between near-infrared and red reflectance of the land surface, normalized by the sum of the reflectances, and is indicative of photosynthetic activity (Myneni et al 1997). The NDVIg data set covers the period 1982–2008 at its 0.07° native spatial resolution and in generating these data explicit corrections for effects of orbital drift and stratospheric aerosols from volcanic eruptions were applied (Tucker et al 2005). To estimate the timing of spring onset, we obtained a newly available long-term (1979–2008) daily freeze–thaw (FT) record based on satellite microwave measurements at ∼25 km spatial resolution (Kim et al 2012). For estimating the primary date of thaw (FTThaw), we applied the 80% rule (12 days having thaw status in consecutive 15 day intervals) as in (Kim et al 2012). At high northern latitudes, FT data have been shown to be closely linked to length of growing seasons (Kimball et al 2004) as well as timing of snowmelt (Rawlins et al 2005). Monthly actual evapotranspiration (AET) and gross primary productivity (GPP) data at 0.5° spatial resolution for 1982–2008 stem from upscaled carbon and water flux observations based on the global FLUXNET tower network (Jung et al 2011). Hereby, the original AET and GPP data have been produced with inputs from climate data as well as satellite vegetation data based on different sensors and satellite platforms. For this study, we recomputed the AET and GPP data for our study period (1982–2008) with inputs from a single satellite vegetation data set (NDVIg) to avoid artifacts of satellite/sensor transitions in the upscaled carbon and water flux data. Finally, monthly climate data (temperature, precipitation and potential evapotranspiration) at 0.5° spatial resolution for the study period (1982–2008) were obtained from CRU (New et al 2002). Precipitation data in the northern high latitudes are relatively uncertain due to sparse and often discontinuous gauge networks. To assess the robustness of corresponding pattern, we utilized an additional precipitation data set from the University of Delaware (UDel), also available at 0.5° spatial resolution (Matsuura and Willmott 2011).
For this study, NA boreal forests include all regions between 50° N–70° N with a tree cover >30%, based on the 2001 MODIS Vegetation Continuous Field metric (Hansen et al 2003). This 'satellite' definition of boreal forests thus includes a relatively small area of more temperate forests in the southwestern portion (figure 1). A focus on more closed canopies reduces the contribution of understory vegetation in the satellite NDVI. We use the MODIS land cover (Friedl et al 2002) and EPA Level II ecozone (CEC 1997) classification to describe patterns (figure 1). All satellite (vegetation, freeze–thaw), upscaled ecosystem flux and climate fields were aggregated (pixel aggregate)/downscaled (nearest neighbor) to a common 0.25° spatial grid on which all statistical analyses were performed. In the case of monthly NDVIg, the mean was taken from the original bimonthly maximum composites and to minimize effects of snow (characterized by negative NDVI values) in the aggregated NDVI data, only grid cells with at least NDVI >0.1 were included in the analysis.
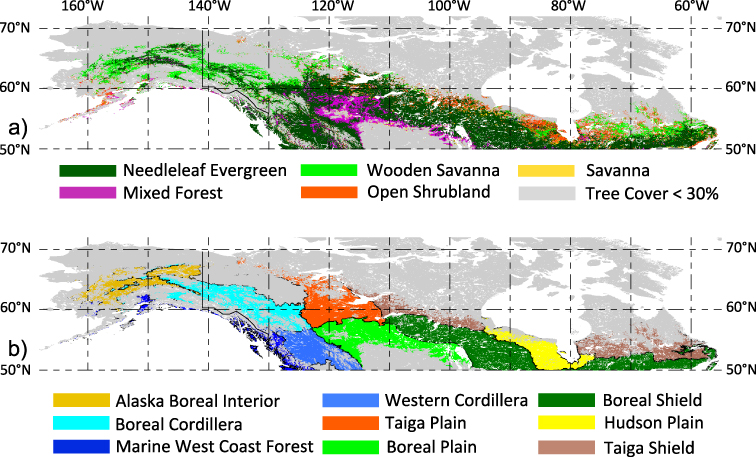
Figure 1. NA boreal forest land cover and ecozones. (a) NA boreal forest land cover based on the MODIS classification scheme (Friedl et al 2002), and (b) EPA level II ecozones (CEC 1997). For (a) and (b), land cover and ecozones are only shown for relatively dense forested regions (tree cover >30% based in the 2001 MODIS vegetation continuous fields (Hansen et al 2003)).
Download figure:
Standard image High-resolution imageIn cases when original time series contain trends, interannual relationships are typically investigated through removal of such trends prior to further investigation. In regards to satellite and climate records, however, extracting corresponding signals is a non-trivial task since commonly used 'detrending' methods based on linear regression can introduce errors if trends are stochastic (Zhou et al 2001). Here, we assess the robustness of interannual relationships by comparing two methods of detrending: (i) first differences in the original time series, and (ii) removing trends based on least squares linear regression in the original time series (Zhou et al 2001). Hereby, we place more emphasis on the first difference method since this method makes no assumption about trends in the original data. Further, our study period is relatively long (nearly three decades) and removal of linear trends over this period may not effectively isolate the desired year-to-year signal. Student's t-tests are used throughout to evaluate statistical significance of correlations. For long-term trend analysis, we also apply least squares linear regression, and use the Vogelsang test (Vogelsang 1998) to assess the robustness of the linear trends. The Vogelsang test can be applied regardless of stationarity in the original time series and also prevents autocorrelation in the time series, or abrupt disturbance events, from generating artificial trends (Beck and Goetz 2011).
3. Results
3.1. Linkages between spring onset and forest greenness at interannual time scales
In a first step, we explored how year-to-year variations in spring onset influence vegetation pattern during the course of the growing season across NA boreal forests. The corresponding correlation pattern, based on first-differenced time series of the original data (see section 2), document how forest greenness (expressed through the NDVI) during May benefits from earlier spring arrivals (negative correlations) almost uniformly across the biome (figure 2(a)). It should be noted that a contribution to the May NDVI enhancement might also arise from diminished snow cover (especially at sub-pixel levels). During June, however, this initial benefit is already greatly reduced over large portions of the NA boreal forests and over some regions in the continental interior, including the Taiga Plain and Boreal Plain, earlier spring arrival is now associated with diminished greenness (positive correlation, figure 2(b)). During July and August, these adverse impacts on vegetation greenness become persistent and spatially extensive across the western and central portion of the NA boreal forest (figure 2(c) and (d)), whereas in the most eastern regions (eastern portion of the Boreal Shield and Taiga Shield) associated vegetation greenness continues to benefit from earlier springs. In August (the month of peak influence) about 28% of the relatively dense NA boreal forests exhibit greenness declines associated with early springs (P < 0.1) suggesting a large-scale phenomenon. During September, signs of negative impacts on vegetation greenness associated with early springs are still evident over the western and central regions (figure 1(e)). A link between earlier springs and adverse summer vegetation greenness has been noted earlier (Kim et al 2012), but the corresponding pattern were much more localized in NA boreal forests possibly a result of mixing associated beneficial and adverse effects on greenness in early and late summer, respectively. The overall patterns associated with early spring onset and adverse peak summer forest greening are regionally more coherent over the western and central climatologically drier portions of the NA boreal forest, but do extend relatively far to the east including the Hudson Plain (figure 1(c) and (d)) with more intermediate moisture levels (figure S1 in supporting information available at stacks.iop.org/ERL/8/024027/mmedia).
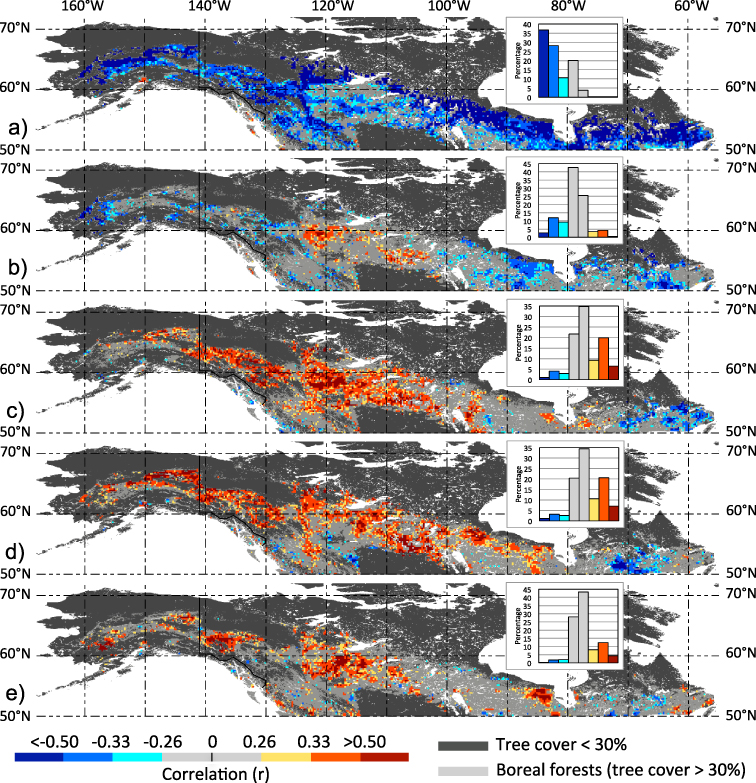
Figure 2. Influence of spring onset on vegetation greenness during the course of the growing season in NA boreal forests. Maps show grid point correlations for the period 1982–2008 between annual timing of spring onset (expressed through FTThaw) and monthly NDVI for (a) May, (b) June, (c) July, (d) August and (e) September, respectively. At each grid point, first differences based on the original time series were computed prior correlations. Absolute r-value categories correspond to significance levels P = 0.2 (r = 0.26), P = 0.1 (r = 0.33) and P = 0.01 (r = 0.50). For each map, frequency histograms showing areal proportions (%) of corresponding positive and negative correlations are provided (see inset panels).
Download figure:
Standard image High-resolution image3.2. Spring onset in relation to other climatic influences on summer forest greenness
Next, we explored how the spring onset influence on NA boreal forest greenness during peak summer (Jul–Aug) compares with other climatic forcings. Surface air temperatures during summer (JJA) are positively associated with peak summer greenness over the eastern regions (including Boreal Shield, Taiga Shield and Hudson Plain) and more localized over the Western Cordillera (figure 3(a)), consistent with temperature-limiting constraints on plant growth in these sub-regions (Nemani et al 2003). Summer (JJA) rainfall rates are negatively linked to peak summer NDVI over the eastern portion of the Boreal Shield and the western Rockies (Western Cordillera and Boreal Cordillera), whereas relatively small clusters in western and central ecozones (Alaska Boreal Interior, Taiga Plains and Hudson Plains) show positive associations (figure 3(b)). The negative associations between peak summer greenness and summer precipitation (and cloudiness) is also consistent with the notion of co-limiting effects of solar radiation on plant growth (Nemani et al 2003), whereas positive associations suggest water supply is limiting plant growth. An index of climatic water deficit in peak summer, the cumulative difference between actual evapotranspiration (AET) based on upscaled water flux observations (see section 2) and potential evapotranspiration (PET; an expression of atmospheric moisture demand) over the period January through August, shows also spatially extensive (positive) associations with peak summer greenness over regions in the western and central NA boreal forests confirming the overall drought sensitivity of these climatologically drier ecozones (figure 3(c)). Cold season precipitation and the depth of snow packs may also influence peak summer vegetation activity especially in regions at higher elevation where summer moisture budgets are more influenced by contributions from spring snow melt (Trujillo et al 2012). Consistent with this assertion, winter (Oct–May) precipitation rates are positively linked with peak summer NDVI over portions of the Western Cordillera and Boreal Cordillera boreal forest regions (figure 3(d)). However, such positive associations are not restricted to higher altitudes but also stretch through the central and eastern ecozones including the Taiga Plain, Boreal Plain, Hudson Plain and Boreal Shield. A comparison of the spatial correlation pattern associated with these warm and cold season climatic influences on peak summer vegetation greenness (figure 3(a)–(d)) with those capturing the influence of the timing of spring onset (figure 3(e)) shows that the latter is a dominant factor in the drier western and central portions of the NA boreal forests. Cross-correlations amongst a subset of these climate predictors are evident, especially for factors that rely on overlapping climatic information (table S1 available at stacks.iop.org/ERL/8/024027/mmedia). However, the markedly different pattern in the corresponding correlations associated with these climate factors suggests that they capture a significant portion of independent influences on peak summer forest greenness. Further, the identified year-to-year correlation pattern exhibit some sensitivity in respect to how the original data were detrended in a prior step and in the case of more uncertain precipitation also depend on data sources (table 1 and figure S2 available at stacks.iop.org/ERL/8/024027/mmedia). But at the level of the entire biome the influence of the timing of spring onset on peak summer greenness is comparable to those related to summer temperature and winter precipitation (table 1).
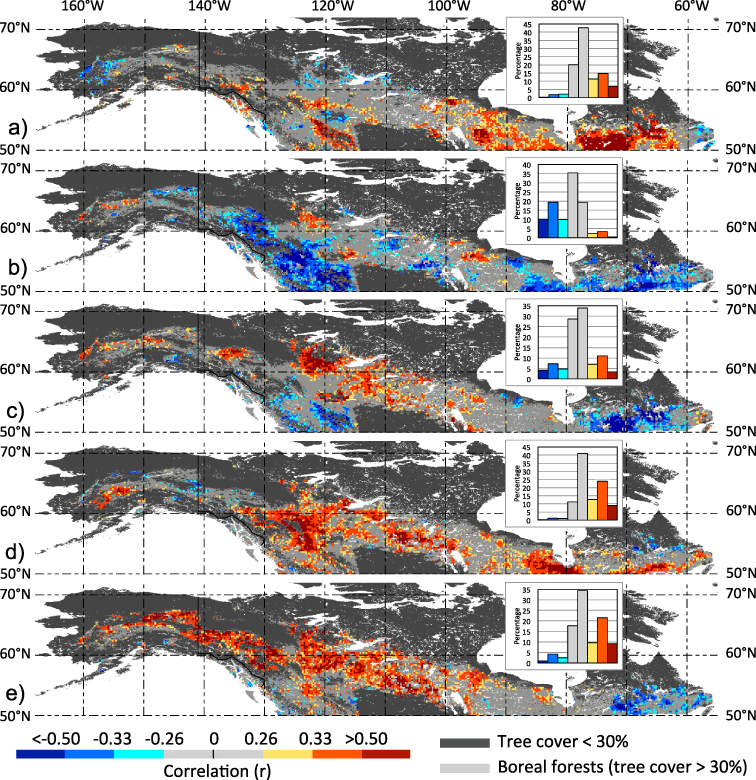
Figure 3. Warm and cold season climatic influences on peak summer vegetation greenness in NA boreal forests. Maps depict grid point correlations for the period 1982–2008 between annual peak summer (Jul–Aug) NDVI and (a) summer (JJA) temperature, (b) summer (JJA) precipitation based on UDel original data, (c) cumulative AET minus PET (Jan–Aug), (d) winter (Oct–May) precipitation based on UDel and (e) spring onset (FTThaw), respectively. At each grid point, first differences based on the original time series were computed prior correlations. Absolute r-value categories are defined in figure 2, and the frequency histogram for areal proportions (%) of corresponding correlations are also shown (inset panels).
Download figure:
Standard image High-resolution imageTable 1. Warm and cold season climatic influences on peak summer vegetation greenness in NA boreal forests. Areal proportions (%) of statistically significant (P < 0.1) correlations between selected climate variables and peak summer (Jul–Aug) NDVI, for the period 1982–2008. Grid–cell correlations were computed on detrended original data using two methods, first differences and removal of linear trends.
| Climate variables | First differences | Removal of linear trend | ||
|---|---|---|---|---|
| Negative (%)a | Positive (%) | Negative (%) | Positive (%) | |
| TemperatureJJA | 2.0b | 21.8 | 0.6 | 33.6 |
| PrecipitationJJA | 29.6 (25.6)c | 3.7 (3.0) | 24.4 (23.3) | 5.8 (3.4) |
| AET–PETJan-Augd | 11.4 | 14.5 | 8.6 | 17.2 |
| PREC–PETJan-Augd | 8.4 (16.6) | 7.9 (6.1) | 8.8 (16.9) | 7.6 (5.6) |
| PrecipitationOct−May | 1.3 (1.2) | 33.0 (20.2) | 0.6 (0.6) | 28.1 (19.5) |
| Spring onset (FTThaw) | 5.2 | 30.9 | 2.5 | 21.8 |
aAreal proportions are based on the total NA boreal forest area (∼3.24 million km2). bStatistical significance thresholds for Pearson correlations are r = 0.33 (p = 0.1, n = 26) and r = 0.32 (P = 0.1, n = 27) for first differences and linearly detrended, respectively. cFor all climate metrics that include precipitation, two data sets (UDel and CRU) are used, whereby all values in brackets are based on CRU data. dThe two indices of climatic water deficit in peak summer, AET-PETJan−Aug and precipitation (PREC)-PETJan−Aug, are formed through summing the monthly differences between the respective variables over the specified time period; both indices assume more negative values under drier conditions.
3.3. Exploration of the 'drought' mechanism associated with timing of spring onset
Is there evidence that shows that the adverse large-scale impact of earlier spring arrival on peak summer vegetation greenness is indeed due to moisture limitations? To investigate this further, we correlated the timing of spring onset with peak summer (Jul–Aug) AET (figure 4(a)). The corresponding spatial (positive) correlation patterns coincide partially with those based on spring onset-peak summer greenness linkages over the western (especially Boreal Cordillera) and central sections of the NA boreal forests (see figure 3(e)). Thus, despite the influence of summer climate, peak summer AET is consistently lower in years with early spring arrival suggestive of water limitations. A similar relationship was found earlier over western NA boreal regions utilizing an AET data set that was generated through a more physically based approach (Zhang et al 2011). In both cases, however, satellite vegetation records were used in the generation of the AET data and, hence, these results are not entirely independent evidence of moisture limitations on plant growth. In fact, independent verification of summer drought related to the spring onset mechanism remains a challenge because for the boreal forest study domain and time frame considered no direct observations of soil moisture are currently available. Further, the two moisture availability indices investigated here (cumulative AET minus PET and PREC minus PET; see table 1) do not appear to capture a summer drought signal related to the spring onset mechanism (figure S3 available at stacks.iop.org/ERL/8/024027/mmedia). This is not surprising since most of the commonly used drought indices either omit entirely or only inadequately incorporate information on seasonal changes in vegetation growth and runoff (e.g. Vicente-Serrano et al 2010).
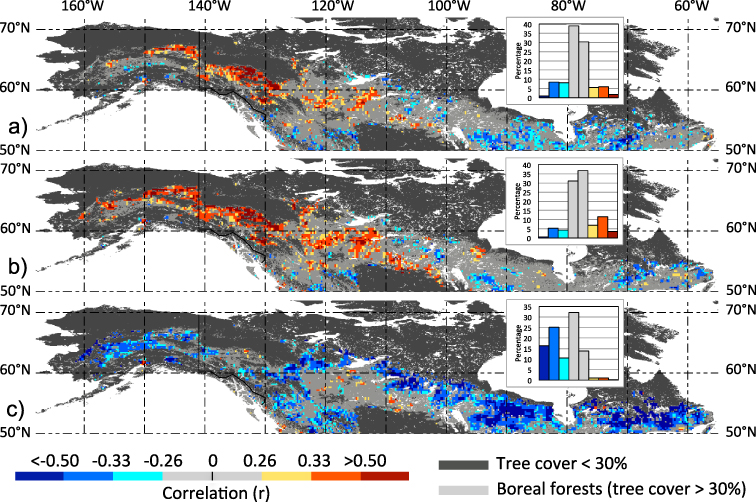
Figure 4. Influence of spring onset on hydrology and ecosystem productivity in NA boreal forests. Maps show grid point correlations for 1982–2008 between annual timing of spring onset (FTThaw) and (a) peak summer (Jul–Aug) AET, (b) peak summer (Jul–Aug) GPP and (c) growing season (May–Sep) GPP, respectively. At each grid point, first differences of the original time series were computed prior correlations. Absolute r-value categories are defined in figure 2, and the frequency histogram for areal proportions (%) of corresponding correlations are also shown (inset panels).
Download figure:
Standard image High-resolution image3.4. Linkages between spring onset and forest productivity at interannual time scales
In a next step, we explored how year-to-year variations in spring onset impact ecosystem productivity across the NA boreal forests. Here, we make use of newly available gross primary productivity (GPP) fields that are based on upscaled carbon flux observations (see section 2). Large portions within the five most western ecozones (Alaska Boreal Interior, Boreal Cordillera, Western Cordillera, Taiga Plain and Boreal Plain) show peak summer (Jul–Aug) GPP declines associated with early spring arrivals (positive correlations), whereas a few relatively small clusters in the eastern and western portions of the Boreal Shield show corresponding GPP increases (negative correlations) (figure 4(b)). These correlation patterns are generally consistent with those linking spring onset and peak summer greenness (figure 3(e)), but are less spatially coherent due to the influence of summer climate on peak summer GPP. Ecosystem productivity over the entire growing season (May–Sep) is generally benefiting from earlier spring onsets (widespread negative correlations; figure 4(c)). But over the regions within the western and central portions (specifically the Boreal Cordillera, Boreal Plain and Taiga Plain), where the corresponding adverse impact on peak summer GPP is large (figure 4(b)), the positive influence of earlier spring arrivals on growing season GPP is effectively cancelled.
3.5. Decadal trends in spring onset and boreal forest productivity
A long-term trend analysis reveals robust shifts to earlier springs within the 1982–2008 study period over portions of the western NA boreal forests that coincide in part with declining trends in peak summer (Jul–Aug) GPP (figure 5(a) and (b)). These trend patterns are also co-located broadly with the corresponding year-to-year (positive) correlation pattern between these variables (figure 4(b)). This consistency in the 'spring onset-peak summer productivity' link at both interannual and longer-term time scales may suggest that shifts towards earlier springs over portions of the western NA boreal forests, specifically over Alaska and adjacent western Canada, may have contributed to a declining trend in peak summer productivity during the last three decades. A closer look from a regional perspective for western NA boreal forests suggests that from the mid to late 1980s onwards spring arrivals shifted continuously to earlier dates and peak summer GPP started to persistently decline (figure 5(c)). However, since the late 1990s no further long-term shift towards earlier springs is evident and this may indicate a diminished influence of this factor on peak summer GPP during the last decade. Alternatively, the pronounced shift towards earlier springs in the 80s and 90s may have crossed an ecosystem threshold and altered the hydrological regime to an extent that summer droughts are much more likely with sustained adverse impacts on summer GPP.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 5. Long-term trends in spring arrival and peak summer GPP in NA boreal forests. Maps depict trends in (a) timing of spring onset (FTThaw) and (b) peak summer (Jul–Aug) GPP for 1982–2008. In (a) and (b), two significance levels for the trends based on linear regression are defined: strong increasing/decreasing trends are those that pass the rigorous Vogelsang test at P < 0.1 (see section 2), and weak increasing/decreasing trends are those where the total change for the 27 year period (e.g. days per 27 years) is equal or larger than the historical standard deviation (based on 1982–2008). In (c), anomalies in spatial averages of timing of spring onset (FTThaw, red) and peak summer (Jul–Aug) GPP (green) for the western NA boreal forests are shown. Plotted are both annual means (thin lines) and corresponding smoothed time series (thick lines) based on a 2nd order Butterworth filter with a five year smoothing time scale. Anomalies are relative to the study period 1982–2008 and negative anomalies in spring onset correspond to earlier springs. Spatial averages for the western NA boreal forests correspond to the area within 130° W–165° W and 60° N–70° N having tree cover >30% (see rectangle in panel (b)). Correlations based on the original time series and after first differencing them (in brackets) are shown. * and ** indicate statistical significance of correlations at P < 0.05 and P < 0.01 levels, respectively.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
Our key finding on the dominant adverse effects of earlier springs on peak summer vegetation greenness, actual evapotranspiration and gross primary productivity at interannual time scales in the western and central NA boreal forest is consistent with the hypothesized reduction in peak summer soil moisture status resulting from water losses (via increases in evapotranspiration, runoff and snow sublimation) in the early part of the growing season (Barnett et al 2005, Jepsen et al 2012). The corresponding negative impacts of earlier springs on late season ecosystem productivity appear to be strong enough to cancel associated productivity gains in the early part of the growing season. A similar 'cancellation' effect has been reported in a previous study on northern ecosystems during the 1990s, whereby ecosystem processes in early and late season were thought to be independent (Angert et al 2005). One important new aspect here is that, for large portions of the NA boreal forests, shifts in spring onsets provide a mechanism through which these early and late season ecosystem processes are coupled.
Several alternative factors may have also played a role in the persistent and spatially extensive (positive) linkages between timing of spring onset and peak summer greenness and productivity in the NA boreal forest. First, local manipulation studies showed that an early spring (and decreased snow cover) exposes the soil to freezing which leads to increased fine root mortality and decreased nutrient availability that in turn adversely impact vegetation productivity later in the growing season (Groffman et al 2001). Second, herbaceous species in the understory of boreal forests may have a fixed lifespan and earlier growth (from earlier springs) would result in earlier senescence (Chabot and Hicks 1982). However, if these factors would be important in the observed spring onset-peak summer greenness links one would not expect corresponding patterns that are sharply centered over the climatologically drier western and central portions of the boreal forest (figures 3(e) and S1 available at stacks.iop.org/ERL/8/024027/mmedia).
A potentially significant contribution to the observed spring onset-peak summer greenness linkages, though, may arise from large-scale climate oscillations that produce seasonally co-varying pattern. For example, warm El Niño–Southern Oscillation (ENSO) years are characterized by both warmer (and earlier) springs and relatively dry summers over large portions of the western and central NA boreal forests (Buermann et al 2003, Shabbar and Skinner 2004), whereby the ENSO signal is thought to be transmitted via an 'atmospheric bridge' (Lau and Nath 1994). Spatially explicit correlation pattern between an ENSO index (Niño1+2) and peak summer NDVI confirm this influence over the western portion of NA boreal forests (figure S4(a) available at stacks.iop.org/ERL/8/024027/mmedia). However, spatially explicit correlations between annual timing of spring onset and the Niño1+2 index are generally much weaker and more localized over the Western Cordillera (figure S4(b) available at stacks.iop.org/ERL/8/024027/mmedia). This suggests that a large portion of the variability in timing of spring onset across NA boreal forests within our study period is not linked to ENSO and, hence, corresponding ENSO influences do not appear to have a large effect on our key findings. However, since strong warm ENSO years are generally associated with warmer and more productive springs over western NA boreal forests (Buermann et al 2003), the results from this study may suggest an additional pathway through which the predominantly winter ENSO signal influences NA boreal summer climates namely also via a 'surface bridge' involving hydrological and biological processes. Other large-scale modes of natural climate variability such as the Arctic Oscillation (AO) and the Pacific Decadal Oscillation (PDO) have also been shown to strongly influence temperature and moisture constraints on productivity pattern in NA boreal regions (Zhang et al 2007), but their temporal characteristics (AO being primarily a winter phenomena and the PDO having little power at year-to-year time scales) suggest no corresponding first order influence on the observed links between spring onset and summer greenness at interannual time scales.
Several field-based and satellite studies have reported declining decadal trends in summer productivity over boreal forests, especially from the mid 1990s onwards, whereby shifts to warmer and drier summer conditions have been invoked as possible explanations (e.g. Angert et al 2005, Beck and Goetz 2011, Ma et al 2012). Our results indicate that the recent decadal shifts towards earlier springs and associated drying in the middle of the growing season over western NA boreal forests may have contributed to these declining trends in summer productivity and may also have played a role in the observed increasing frequency of large fire years and late season (Aug–Oct) fires during the 1980/90s (Kasischke and Turetsky 2006). Further, these decadal linkages between spring productivity gains and summer productivity losses may also have a been a contributing factor, in addition to warming-induced increased autumn respiration, in the recent coordinated shifts in the seasonal cycle of atmospheric CO2 as observed at some high-latitude monitoring stations (parallel shifts towards earlier 'downward zero crossing' and 'upward zero crossing ' dates in spring and autumn, respectively) (Piao et al 2008).
5. Conclusion
At the biome level, we find that the amount of winter precipitation and associated depth and extent of snow packs by itself has a large positive influence on NA boreal forest greenness in the later portion of the growing season (e.g. figure 3(d) and table 1), consistent with earlier results (Grippa et al 2005). Further, the timing of spring onset in NA boreal forests is predominantly influenced by spring temperatures and at more regional scales also by the amount of winter precipitation (see figure S5 available at stacks.iop.org/ERL/8/024027/mmedia). Taken together, these results have severe implications for the boreal ecosystems under future climate projections. It is quite conceivable that the 'spring onset-peak summer productivity' mechanism will grow in importance with the projected accelerated warming (and persistent shifts to earlier springs) in the northern high latitudes (Solomon et al 2007). The projected increases in predominantly cold season precipitation over these regions (Solomon et al 2007) will have only limited buffering capacities in respect to summer drying since portions of this increased water supply will be lost via increased early runoff and longer evaporative periods resulting from earlier springs. In addition, a greater proportion of cold season precipitation will fall as rain versus snowfall with the effect of further increasing early season runoff and also advancing spring arrival towards earlier dates. Lower soil moisture status and higher suboptimal temperatures during summer (Barber et al 2000) may lead to persistent adverse impacts on boreal forest growth and survival (Hogg et al 2008) and may accelerate the disturbance dynamics (Kasischke and Turetsky 2006, Kurz et al 2008), setting the stage for initiating large-scale boreal forest dieback.
Acknowledgments
Funding for this study was provided by the National Aeronautics and Space Administration Carbon Cycle Science Program (Grant: NNX11AD45G). We thank E H Hogg, N P Molotch, and M P Girardin for constructive comments. We also thank P S A Beck and S J Goetz for providing the computer program to run the Vogelsang test.