
Abstract
The endoskeleton of echinoderms (Deuterostomia: Echinodermata) is of mesodermal origin and consists of cells, organic components, as well as an inorganic mineral matrix. The echinoderm skeleton forms a complex lattice-system, which represents a model structure for naturally inspired engineering in terms of construction, mechanical behaviour and functional design. The sea urchin (Echinodermata: Echinoidea) endoskeleton consists of three main structural components: test, dental apparatus and accessory appendages. Although, all parts of the echinoid skeleton consist of the same basic material, their microstructure displays a great potential in meeting several mechanical needs according to a direct and clear structure–function relationship. This versatility has allowed the echinoid skeleton to adapt to different activities such as structural support, defence, feeding, burrowing and cleaning. Although, constrained by energy and resource efficiency, many of the structures found in the echinoid skeleton are optimized in terms of functional performances. Therefore, these structures can be used as role models for bio-inspired solutions in various industrial sectors such as building constructions, robotics, biomedical and material engineering. The present review provides an overview of previous mechanical and biomimetic research on the echinoid endoskeleton, describing the current state of knowledge and providing a reference for future studies.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Introduction
Sea urchins (Echinodermata: Echinoidea) are known to have been in existence since the Middle Ordovician, about 460 million years ago [1]. During the Early Jurassic, they underwent an intensive adaptive radiation leading to a variety of specialized forms and lifestyles adapted to different marine habitats [2–13]. Echinoids are traditionally subdivided into two groups: regularia and irregularia, mainly identifiable based on test morphology and lifestyle [14, 15]. Regular echinoids are typically spherical in shape with the peristome (mouth region) on the central oral side and the periproct (anal region) aborally located. The area spanning from the apical system throughout the peristome is divided in five ambulacral and five interambulacral fields, each one characterized by ten double columns of different skeletal plates with species-specific fine-relief ornaments [16]. Regular echinoids possess a prominent pentaradial symmetry superimposed on the ancestral echinoderm bilateral symmetry. In contrast, irregular echinoids are typically aboral-orally flattened and elongated or heart-shaped. The peristome is orally located, but not necessarily in the centre of the oral surface. The periproct migrated from the central aboral side towards the oral side assuming variable positions in the test [12, 17]. The ambulacral fields are often restricted to the aboral side forming the petalodium [18]. Thus, irregular echinoids typically possess a strong bilateral symmetry superimposed on the radial symmetry acquired [4–8, 19–25]. The evolutionary success of echinoids is undoubtedly due to the strategic employment of their endoskeleton, macroscopically consisting of three main functional components: test, dental apparatus (Aristotle's lantern) and accessory appendages (such as spines and pedicellariae) [26, 27] (figure 1).
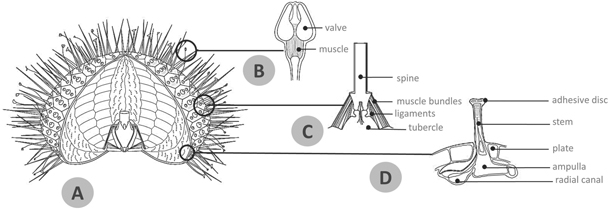
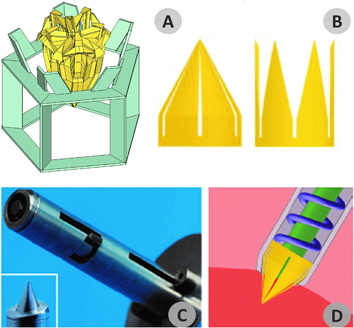
Figure 1. Overview of the echinoid test and appendages. (A) Schematic reconstruction of regular echinoid test showing its overall external and internal anatomy including the lantern system. (B) Detail of pedicellaria tip illustrating the three valves and muscles. (C) Detail of the spine-test articulation showing the typical ball-and-socket joint in a vertical section: the spine is articulated to the plate tubercle through the muscle bundles and ligaments of the 'catch apparatus'. (D) Detail of a tube foot displaying the main components of its microanatomy: basal ampulla, stem and apical adhesive disc including tiny skeletal ossicles.
Download figure:
Standard image High-resolution imageIn the course of evolution, the echinoid skeletal parts transformed in morphology and physiology adapting to novel functions [28]. For example, in some species the main function of the spines shifted from protection to burrowing [29–33]. Also, pedicellariae, the small pincer-like appendages, developed different forms, including venomous types [34]. In addition, the morphology of the dental apparatus differed due to feeding strategies, such as scraping and crushing, or it has been entirely eliminated [35–38]. Complementarily, the modifications during evolution have specialized and adapted these skeletal parts to efficiently absolve specific mechanical roles. In particular, spines and test protect the animal by withstanding biotic (e.g. predatory attacks) and abiotic (e.g. strong wave motion or substrate impact during burrowing or locomotion) mechanical stresses [39–46]; pedicellariae, provide further defence and are used for cleaning the echinoid's epidermis [34, 47]. Aristotle's lantern plays a direct role in multiple activities such as gripping, scraping, digging, and even locomotion [48]. In particular, the lantern, which consists of an integrated system of 40 skeletal elements, joined and moved by specific muscles and ligaments, represents one of the most complex and optimized biomechanical models in the animal kingdom [49–53].
Due to its unique features, it does not surprize that the constructional design of the echinoid skeleton has attracted the interest of both biologists and engineers. Accordingly, mechanical engineering and material science principles, methods and tools have been applied in exploring the mechanical performances of sea urchins as an integrated system or single component [23, 45, 46, 54–63]. This biomechanical approach provided important biological insights on form-function skeletal features, taxa comparisons, ecological and evolutionary trends and adaptive meanings, as well as, new functional principles used to design innovative bioinspired technical solutions [27, 46, 54, 64–68]. Echinoid skeletal components are structurally and functionally organized regarding, among others, lightness, stability, strength, flexibility and stress resistance. Presently, due to the availability of novel analytical methods, the underlying principles can be better understood and transferred into building constructions and industrial products; a process known as 'biomimetics' and 'bionics' [69]. Otto Herbert Schmitt an engineer and physicist coined the term biomimetics in 1957 and its approach was regulated and certified in 2015 by the International Organization for Standardization (ISO 18458) [70]; whereas, the term bionics, a combination of the words 'biology' and 'technics', was coined by the US Air Force Major J E Steele in 1960 [71–75]. Often used as an equivalent, both terms identify a design process inspired by nature that generated innovative technological solutions. Over the past decade, other terms have occurred in conjunction with this process, such as biomimicry, biomimesis, bio-inspiration, nature-based solutions, biologically inspired designs and numerous others; although often used as synonyms, each one differs in objectives, principles and approaches [69, 74–76].
The present review provides an overview of recent knowledge on echinoid skeletal structures. Its intention is to identify the main morphological features and mechanical aspects, in order to provide a reference for future research on biomimetic applications. Accordingly, the following issues will be discussed in detail: (1) current knowledge of biomineralization and material properties of the echinoid endoskeleton; (2) skeletal microstructure (stereom); (3) the three main skeletal components: i.e. test, Aristotle's lantern and accessory appendages; (4) biomimetic process and echinoid-inspired applications in building constructions, robotics, biomedical and material engineering.
1. Biomineralization
The biomineralization process in echinoid skeletons has been extensively investigated throughout different developmental stages from larvae to adults [77–85]. Detailed mineralogical analyses revealed that its mineral matrix consists of calcite, containing up to 15% magnesium [86, 87]. Hence, the echinoid skeleton is generally considered a high-magnesium calcitic structure, although its magnesium content can vary significantly according to species and specific skeletal parts, as well as, environmental factors such as temperature or pH [88–92]. These variations determine different mechanical properties of the skeletal parts [88, 93]. The calcite in echinoid skeletons displays the optical behaviour of a monocrystalline structure with definite orientation of the optical axes [87, 94]. In terms of mechanical behaviour, the rupture response of the echinoid biocalcite results in conchoidal fractured surfaces, which differ from the well-defined cleavage of pure calcite crystals [95–98]. For many years, this fracturing behaviour was attributed to the presence of organic components (proteins) within the stereom structure [95]. Seto et al (2012) later found evidence that this behaviour is mostly due to the particular echinoid calcite structure [99]. Indeed, this calcite is a mesocrystal composed by numerous aligned calcite nanocrystals (∼100 nm) embedded in a matrix of amorphous calcium carbonate (ACC) and macromolecules [95–102]. These last two components cause the conchoidal fracture properties. Echinoid biocalcite has often been discussed as representing a composite material because it contains up to 0.2% proteins by weight [84, 103, 104]. From a material engineering perspective, materials composed of two or more constituents with different physical, chemical and mechanical properties are defined composites. The combination of different constituents produces a material with advantageous emerging properties, strongly different from the properties of the same constituents [45]. This is usually the case when the fraction of each composite reaches a relevant amount of the total volume [105]. When the amount of one of the components is too low, the material is not considered a composite; in fact, the second constituent affects the material properties by interacting with the main component rather than contributing its own advantageous properties to the material composition [105].
Vertebrate bone for example, represents a high-performance composite material consisting primarily of collagen and hydroxyapatite. The mineral component provides bone with mechanical rigidity and load-bearing strength, whereas the organic fibrous component provides elasticity and flexibility [106]. In quantitative terms, the hydroxyapatite fraction should account for 35% of the volume in order to reinforce the skeletal material effectively. Nevertheless, the amount of hydroxyapatite in vertebrate bone reaches roughly 50% and the collagen represents the other 50% providing advantageous tensile properties; for this reason, vertebrate bone represents a true composite material, of which anisotropy provides considerable strength and stiffness in at least one direction [45, 106, 107].
Herman et al (1988) demonstrated that the amount of organic matrix in echinoid calcite is not sufficient to form a continuous layer within the stereom and does not represent a considerable amount of the total volume; thus, it cannot be considered as an effective constituent in making the echinoid calcite a composite material [96, 108]. However, Seto et al (2012) demonstrated that the mesocrystal structure of the echinoid calcite contains between 8 and 10 wt % ACC in mature spines, consequently revealing that ACC calcite could itself serve as a second component of this composite material [99].
Composite systems, usually feature the advantage that cleavage propagation is prevented by a suitable alternating arrangement of stiff, strong materials with less stiff materials creating a functional interface where the latter, having a reduced elastic modulus, assumes a stress-breaking role absorbing stresses [105, 107, 109]. This is the case of nacre that is composed of 95% aragonite and 5% proteins and polysaccharides, as well as, vertebrate bone [45, 107]. The employment of calcified collagenous fibres consequently results in an anisotropic material, which is stiff and tough in one direction, but brittle in at least another [45, 107].
Recently, Lauer et al (2020) demonstrated that unlike mechanical properties of other biogenic ceramic composite materials, such as nacre, the combination of high Mg-calcite with ACC and organic phases have little effect on macromechanical properties of the Heterocentrotus mamillatus spines [110]. Thus, although the micromechanical properties of the echinoid skeleton are governed by the interplay of ACC, organic phase and Mg calcite [96, 99, 111], the macromechanical properties seem mainly governed by the porous stereom structure and architecture resulting in a remarkable damage tolerance [110].
Interestingly, the crystallographic design and macromolecule distribution makes the echinoid bio-calcite a more isotropic material [112]. In this regard, it has been demonstrated that the anisotropy is larger in synthetic crystals than in young sea urchin spines; whereas, mature spines have an extended anisotropy, ranging between those of synthetic crystals and young spines, suggesting the existence of remarkable differences in the biological crystal composition during spine formation and growth [113]. In contrast, vertebrate bones (such as femurs) display a clear and defined preferential orientation of collagen and apatite inside trabeculae, as well as, a highly anisotropic trabecular architecture; thus, it is capable of transferring loads more effectively in only one direction [45, 107]. However, apart from the mineral composition, echinoid stereom is similarly characterized by a variable oriented trabecular architecture ensuring a more directional resilience [57, 114]. Moreover, due to its trabecular meshwork, the echinoid stereom is a lightweight construction and possesses a high level of robustness, e.g. allowing the applied forces to bypass malfunctional trabeculae and to be transferred to the functional surrounding ones. [57, 94, 114].
2. Stereom
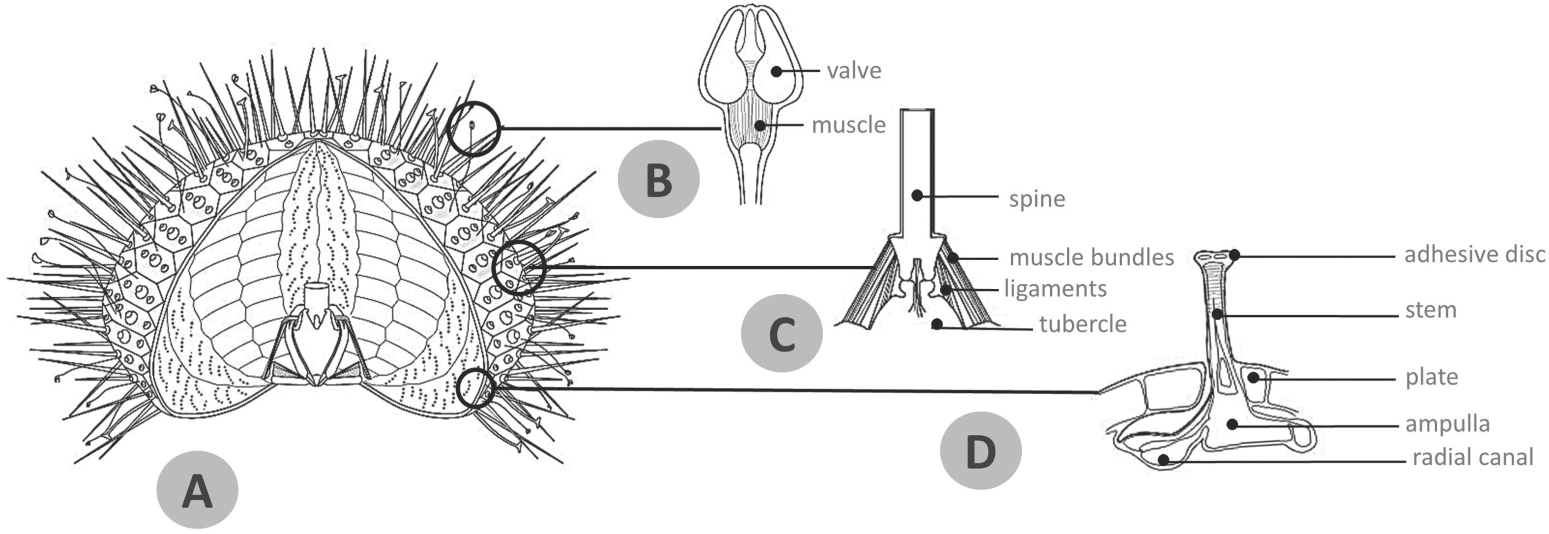
Stereom [115] is a 3D mesh of trabeculae, i.e. struts, made of biocalcite [114]. It represents a key element responding to the principles of robustness, lightness and stability, due to three primary factors: (1) material composition and related mechanical properties based on material variations through strategic substitution of calcium (Ca) with magnesium (Mg) in the calcite crystal, and alterations of fracture behaviour [94–98, 102, 116–121]; (2) high structural porosity of about 50%–75% [45, 59, 122]; (3) large structural variability (related to e.g. phylogeny, functional adaptations, growth rate and soft tissue types) allowing specific mechanical demands to be met [46, 54–59, 84, 114, 123, 124]. Consequently, this lightweight structure denotes an important adaptive achievement within the entire phylum Echinodermata contributing to its evolutionary success [125–127]. The complex constructional design of the stereom varies from species to species and within both individuals and skeletal elements. Nevertheless, known far away in time [128], this structural variability was described in detail by Smith (1980) identifying ten different stereom types in the test: imperforate, microperforate, simple perforate, galleried, rectilinear, retiform, laminar, fascicular, labyrinthic and irregular perforate [114]. All of which can be employed in a number of combinations, creating species-specific 3D structural patterns easily recognizable in scanning electron microscope (SEM) images. Architectural variability and possible modulations based on specific mechanical needs have been described in several studies regarding: (1) the test and its individual plates [46, 55–57, 87, 97, 116, 124, 129, 130]; (2) Aristotle's lantern ossicles [52, 131]; and (3), more frequently, spines [27, 30, 52, 54, 58, 61, 65, 123, 132–135]. As a rule, stereom density tends to increase in regions subjected to high mechanical stresses resulting in imperforate or microperforate types; in particular, this occurs in those areas exposed to high frictional and compressive stresses, such as tubercles (figures 2(C) and (E)), lantern (figure 3(B)), rotulae (figure 3(C)) and spine bases [27, 52]. Also, the microstructure tends to be specialized in regions subjected to directional tensile stresses, i.e. the sutural areas characterized by galleried stereom, which offers a suitable insertion pattern for the attached ligament bundles (figures 2(C)–(E), 3(D) and (E)) [27, 52, 54–57, 114, 123]. On the contrary, labyrinthic stereom is found in zones subjected to multi-directional mechanical stresses, such as in the centre of the plates (figures 2(C)–(E)), lantern demi-pyramid (figures 3(B) and (C)) and in the radiating layer of camarodont spines [52, 54, 114]: according to Grun and Nebelsick (2018) labyrinthic stereom could equally distribute stresses in multiple directions, thus reducing local stresses [46].

Figure 2. SEM micrographs showing microstructure and stereom variability of the test plates. (A) Topographic reference of the interambulacral region inspected. (B) Schematic reconstruction of an interambulacral plate: (a) top view, (b) internal view, (c) lateral view. (C) Top view of Paracentrotus lividus interambulacral plate; (D) enlargement of figure (C). Labyrinthic stereom with variable porous texture is the dominant microstructural pattern. In the wide circular insertion area of the overall catch apparatus it is possible to distinguish specific stereom patterns related to muscle and ligament insertions, density and sizes of porosity, closer and more regular in the ligament area. Adjacent to the tubercle where the stereom structure tends to become imperforate, pore size decreases. (E) Vertical section of the plate showing a high diversity of the stereom microarchitecture according to zones and related specific mechanical needs. (F) Details of stereom types detected: (1) imperforate stereom; (2) labyrinthic stereom; (3) galleried stereom; (4) microperforate stereom. Bar = 100 μm. lia = ligaments insertion area, mia = muscles insertion area, tb = tubercle.
Download figure:
Standard image High-resolution image
Figure 3. SEM micrographs showing structure and stereom variability in the Aristotle's lantern components. (A) Topographic reference of the skeletal parts inspected. (B) Detail of the intrapyramidal suture: galleried stereom is dominant in those areas near the suture running perpendicularly in correspondence to its borders; dense labyrinthic stereom prevails in the unspecialized superficial areas. Bar = 100 μm. (C) Frontal view detail of a jaw showing the epiphyseal arch and the compass-rotula complex. Labyrinthic stereom is the dominant microstructural pattern. At the epiphyseal fossa level, an increased open pattern provides suitable anchorage for the protractor muscles. Bar = 400 μm. (D) Internal view of a demi-auricle showing the attachment area of the retractor muscle. Fascicular-labyrinthic stereom prevails in the upper part of the auricle, whereas microperforate stereom is the dominant pattern in the central region. Bar = 500 μm. (E) In detail the suture of the auricle basal plate (opened complementary parts): galleried stereom runs perpendicular to the sutural surfaces. (F) Vertical section of the auricle basal plate showing the variability and complexity of its internal microarchitecture in relation to the ambulacral pore arrangement: labyrinthic and microperforate stereom prevailing. Bar = 100 μm. irm = retractor muscle insertion area, c = compass, de = demi-epiphysis, dp = demi-pyramid, ef = epiphyseal fossa, p = ambulacral pore, r = rotula, s = suture, subapical fossa, t = tooth.
Download figure:
Standard image High-resolution imageIn the past decades, the mechanical design of the stereom has been extensively studied in a two-dimensional view [16, 33, 84, 114, 129, 130, 134, 136, 137]; however, with the advent of affordable high-resolution computed tomography (CT) scanning, recent studies explored the stereom using 3D modelling reconstruction, 3D topological and structural analysis (e.g. finite element analysis, FEA). These modern methods allow detailed analyses of mechanical properties, lightweight constructions and load-bearing systems [55–57, 59, 124, 135, 138]. Accordingly, different mechanical tests on the skeletal layout demonstrated how these stereom variabilities have diverse structural implications [54, 61, 67, 90, 122, 132, 133, 139].
Hitherto, it is also important to remark that the echinoderm skeleton is a proper mesodermal tissue, and that the living stereom contains an organic stroma, consisting of cells and extracellular matrix including collagen fibres [45, 114]. The stroma significantly contributes to the integrity of the skeleton providing indispensable resistance and flexibility qualities. In general, this organic component: (1) reinforces the endoskeleton, providing greater mechanical resistance to the overall structure and continuity to the related ligaments, thus avoiding the risk of fracture at low applied forces [140]; (2) transforms the test into a flexible jointed integumental layer meaningfully reducing the impact of bending stresses [140, 141]; (3), acting as an energy absorbance system and stress-breaker interrupting the propagation of fractures due to material component discontinuity (stereom + stroma = rigid + elastic components) [52, 142–144]; (4) confers reinforcement, support and potential repair to the mineral structure [95, 103, 108, 145, 146].
3. Test
The echinoid test (figure 4) is a multi-element system consisting of a number of skeletal plates joined by sutures. These sutures can be characterized by the presence of interdigitating articular surfaces (comb-joints) often bound together by short collagenous ligaments [23, 27, 33, 46, 57, 63, 140, 147–149]. This constructional design fulfils several mechanical principles acting as a resistant, lightweight, load-bearing and load-transferring system, as well as being an attachment point for appendages. Structural strength is achieved by hierarchical constructional adaptations, such as: overall shape, plate layout and arrangement (trivalent vertex arrangement, in which three plates meet in one point), skeletal interlocking and reinforcements (e.g. internal buttressing), material distribution and stereom diversity [27, 46, 55–58, 63, 140, 149–151]. These skeletal features have been described as functional strategies which, suitably combined with adaptations of the connective tissue components, allows the echinoid test to withstand compressive, tensile and bending stresses [46, 55, 63, 109, 140, 141, 149]. In particular, collagenous sutural ligaments play a central role in increasing the structural strengthening of the test by binding rigid calcite plates at sutures [140, 141]. By measuring the breaking forces of the Strongylocentrotus purpuratus skeleton with intact or removed soft tissues, Ellers et al (1998) demonstrated that skeletons without ligaments broke at lower apically applied forces in respect to those with ligaments [140]. Different is the case of the minute clypeasteroid Echinocyamus pusillus, of which Grun and Nebelsick (2018) showed that soft tissues do not possess a significant structural function. However, the overall layout and plate connections between Strongylocentrotus and Echinocyamus are fundamentally different due to the extensive skeletal plate connections in the Echinocyamus responsible for its overall stability [46, 55–58, 149]. Some echinoid morphologies are also optimized with respect to hydrodynamic property adaptations such as the lunulae of sand dollars, which are considered to reduce lift when sand dollars are on the sea-floor surface and subjected to strong currents [62].

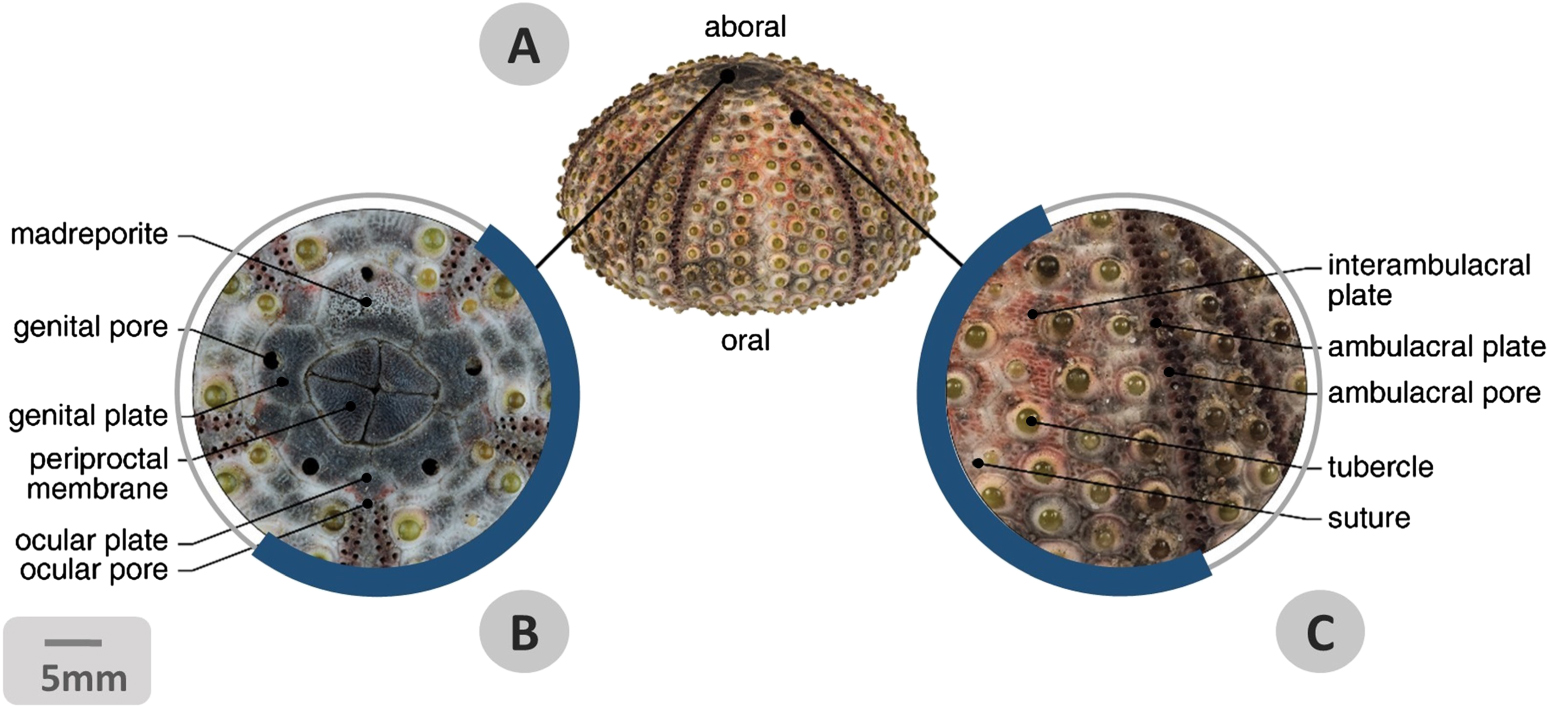
Figure 4. Stereo-micrographs of regular echinoid test. (A) Lateral view of Arbacia lixula test deprived of its soft tissue component and accessory appendages. It is possible to distinguish an 'oral' side where the mouth opens and an 'aboral' side with the anal opening. (B) Pattern of the apical pole: the detail shows the madreporite, the five genital plates with respective genital pores; five ocular plates with pores; the periproctal membrane with the centrally positioned anus. (C) Details of the lateral side of the test showing the arrangement of double columns of interambulacral and ambulacral plates, the latter ones recognizable by the tube-feet pores. All plates display articular tubercles for spines. Central sutures between two adjacent plate columns are visible. Bar = 5 mm.
Download figure:
Standard image High-resolution imageDue to the structural form and architecture of the test, echinoids have been extensively investigated in order to understand their constructional design and mechanical behaviour in detail [23, 45, 46, 55, 60, 63, 68, 107, 148, 150, 152–154]. Detailed morphospace analyses were carried out to explain and predict extinct and extant echinoid test shapes by considering possible phylogenetic, physical and mechanical factors [154–158]. Thompson (1917) in particular, carried out a pioneer study on test shape using a liquid drop analogy to describe the shape and growth of regular echinoids [158]. Ellers (1993) supported this hypothesis using the thin shell theory to explain test curvature defining the echinoid morphospace in two parameters: (1) the apical curvature; (2) a proportion of the vertical gradient of pressure to the internal coelomic pressure [156]. Seilacher (1979) proposed that the echinoid test should be analysed as a mineralized pneu-structure that grows when internal pressure exceeds external tension, varying its morphology through plate growth [23, 28]. However, Ellers and Telford (1992) measured the internal coelomic pressure in the regular sea urchin S. purpuratus and Lytechinus variegatus [159]. They found that internal pressure fluctuates rhythmically about −8 Pa and was negative for 70% of the time, disempowering the pneu-hypothesis that requires an internal positive pressure [23, 28, 160]. These rhythmical fluctuations in pressure could be mainly caused by the lantern movements that change the curvature and tension of the peristomial membrane [159]. Telford (1985) analysed the test as a dome structure utilizing both the membrane theory and static analysis to determine its behaviour under different loads; thereby assessing the hypothesis that the test form was constructed to resist external forces [63]. On the whole, taking into account these and other studies, test form and growth were described and explained using different theoretic models, based on a total of nine hypotheses, in addition to different computational models [for review see 152]. The echinoid test growth is mainly based on two combined processes, namely: plate addition, i.e. the insertion of new plates in the apical system [21], and plate growth, based on a peripherally accretion or reabsorption of skeletal material [161]. However, the main distinctive feature of the growth process lies in the mutable collagenous tissue (MCT) present at plate sutures that can undergo rapid changes in mechanical properties (switching reversibly between stiff and compliant states) accommodating little movement and growth [for review see 162]. In particular, sutures allow growth maintaining a space between plate margins ('plate gapping') [152, 155, 163] in a manner that they do not unite and continuously expand interacting with the adjacent plates. Usually in regular echinoids, sutures remain open up to the adult stage providing the test some degree of flexibility and mechanical advantages in sustaining loads [140, 141].
Modern methods such as 3D acquisition (e.g. µCT and photogrammetry), digital modelling and simulation, e.g. FEA are recently being adopted, providing novel answers to questions about test morphology, functional performance and mechanical behaviour (figure 5) [46, 55, 57, 60, 138, 141, 164, 165]. As pioneers in this field, Philippi and Nachtigall (1996) conducted FEA-analysis describing the behaviour of the regular echinoid test (Echinus esculentus) under diverse loads [60]. Their studies highlighted the structural load-bearing efficiency of the test and interpreted its peculiar spherical shape as the most adapted form to sustain the tensile stresses resulting from the tube feet activity [60]. Recently, Grun and co-workers focussed on the clypeasteroid skeleton using x-ray µCT, SEM observations and physical and virtual tests in order to analyse the hierarchical structural design of the E. pusillus test [46, 55, 57]. They displayed in detail the mechanical properties of the test at different hierarchical levels, i.e. from the overall shape—although consisting in a discontinuous structure divided into several polygonal plates, it behaves as a monolithic structure—to the plate micro-architecture, internal supports and stereom variability, all described as specific functional devices for bearing and transferring loads.
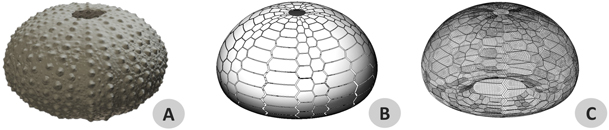
Figure 5. Virtual models of Paracentrotus lividus's test. (A) 3D model obtained with a photogrammetric reconstruction. (B) Parametrized geometry model with visible plates and sutures. (C) 3D mesh used for finite element analyses.
Download figure:
Standard image High-resolution image4. Aristotle's lantern
Most regular echinoids, extant or extinct, possess a complex dental apparatus, traditionally called Aristotle's lantern. The apparatus is a biomechanical and dynamic system arranged according to perfectly pentameral symmetry and consisting of an intrinsic part, the lantern itself, and an extrinsic part, the perignathic girdle, i.e. the inner edge of the test [49–53]. These two parts are connected by muscle bundles (five pairs of retractor muscles and five pairs of protractor muscles), and ligamentous structures (peristomial membrane and five pairs of compass depressor bundles) [51] (figure 6). The lantern consists of forty anatomically distinct skeletal ossicles: ten demi-pyramids, ten epiphyses, five rotulae, ten compasses and five teeth (figure 7) [52]. They are all reciprocally joined by specific articulations (movable joints, semi-movable or rigid sutures), interconnected by articular ligaments and moved by anatomically and functionally well-defined muscles consisting in five pairs of retractor and protractor bundles, five massive inter-pyramidal muscles and five compass elevator muscles. The lantern muscular component is also represented by other muscular elements, namely myocytes of the lantern coelomic epithelium, which are involved to a minor extent (such as the thin muscle layer included in the compass depressor ligaments) [49, 53, 166–169].
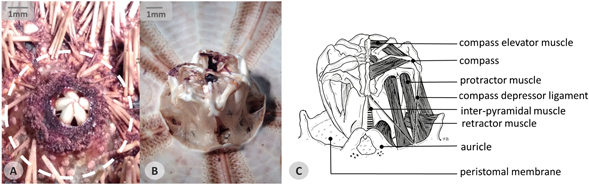
Figure 6. Aristotle's lantern. (A) Stereo-micrograph representing an oral view of a Sphaerechinus granularis sample showing in detail the peristomial membrane (dotted circle), the mouth opening and five teeth tips. (B) Stereo-micrograph of the lantern in a dissected specimen. (C) Schematic illustration of the lantern anatomy showing: five compasses, respective compass elevator muscles and depressor ligaments; protractor and retractor muscles; inter-pyramidal muscles; auricles; peristomal membrane.
Download figure:
Standard image High-resolution image
Figure 7. SEM micrographs of a real Aristotle's lantern (A–C) compared with a three-dimensional reconstruction (D–F). (A) Front/lateral view of the lantern. The preparation method (semi-digestion of soft tissues) emphasizes the skeletal structure and permits observation of the five jaws comprising their different components. (B) Internal view of two isolated pyramids, compass and teeth. The lateral sides of the pyramid possess comb-like indented inner borders and transversely striated surfaces with regular ridges for the inter-pyramidal muscle attachment. (C) Front/lateral view of a tooth in toto showing a T-shaped structure (apical plumula partly removed). Bar = 1 mm. (D) A prospective view of the segmented 3D model of Paracentrotus lividus lantern based on micro-CT imageries. (E) A virtual section of pyramids showing teeth position. (F) Isolated teeth. Blue = pyramids and compass, yellow = teeth. Bar = 1 mm. c = compass, de = demi-epiphysis, dp = demi-pyramid, k = keel, m = midshaft, py = pyramid, pl = plumula, r = rotula, t = tooth, tp = tip.
Download figure:
Standard image High-resolution imageConversely, irregular echinoids do not generally possess a lantern, although in juveniles of Cassiduloida and Spatangoida this can appear as a vestigial trait, with the exceptions of adult Holectypoida and Clypeasteroida, [37, 38]. However, these persistent lanterns differ remarkably from regular lantern models: they are flattened and relatively larger, non-protrusible [38] and provided with teeth that move horizontally with respect to the substrate and designed to crush sediment rather than to grasp [170, 171]. Furthermore, in contrast to the lantern of regular echinoids, these flattened types appear to be used only for feeding: the Aristotle's lantern of regular sea urchins is employed in other important activities [33, 53, 166] such as digging, locomotion, respiration and circulation of coelomic fluid [170, 172, 173].
From Aristotle's studies [174], the unique structure of the echinoid dental apparatus has attracted the attention of many scientists [128, 175–178], but only in the last century the interest of biologists and engineers in the lantern has increased, resulting in a wide range of studies aimed to address its morphology [51–53, 179], physiology [180–182] and biomechanics [49, 49–53, 171, 173, 183–194].
Static and dynamic mechanical studies were carried out on the echinoid lantern, specifically on its skeletal ossicles, muscular system and ligaments [50–53, 194–198], as well as, on the peristomial membrane (figure 6(A)), a flexible area consisting mainly of fibrous connective tissue surrounding the mouth and connecting the lantern to the test; with its dynamic mechanical behaviour it contributes to the lantern's stability and motility [168, 169, 199, 200]. Biomechanical models, experimental mechanical tests and computer simulations were elaborated and integrated to determine lantern movements, muscular forces and constraints during different activities in regular echinoid lanterns [49, 184, 192, 193], whereas other mechanical studies were addressed to define the biting forces developed by the dental apparatus in sand dollars [170]. It was assessed that the overall lantern can show resistance to different mechanical stresses directly or indirectly related to motor activities by means of a number of specific macro- and micro-structural adaptations. From a macrostructural perspective, the first mechanical advantage of the lantern lies in its strategic subdivision into complementary parts and correlated pieces, starting with the five multipiece jaws (figure 7), each consisting of distinct elements sutured together (two symmetrical demi-pyramids and two symmetrical demi-epiphyses) providing a perfect alveolus that contains and protects the long internal tooth ensuring its continuous growth (see below) [52] (figures 7(B) and (C)). The second advantage regards the jaws that are joined to each other by means of complex multivalent articulations endowed with specialized articular ossicles, known as rotulae [52] (figure 3(C)). They play a role in the basic opening and closing of the jaw, modulating its reciprocal tilting and swinging, and in the independent movements of the compasses (raising/lowering) on the aboral side of the lantern. These are sophisticated devices enabling the structure to be mechanically versatile, resistant and deformable [51–53, 143]. Nonetheless, the major complex adaptations were found in both skeletal microstructural variations/differentiations (figure 3) and material composition. The micromechanical design of the skeletal parts of the lantern was extensively investigated and described using SEM by Candia Carnevali and co-workers in comparative studies of the cidarid [51] and camarodont [52] lanterns. Detailed SEM studies also focussed on the micro-structure of sea urchin teeth [171, 183, 187–191, 200]. Subsequently, pyramids and teeth were further analysed employing Micro-CT imaging, which permitted the acquisition of 3D images leading to detailed insights into different species-specific geometries and microstructures [120, 131, 201–205] (figures 7(D)–(F)). These studies demonstrated that the lantern ossicles tend to have a similar basic organization in terms of adaptive stereom variability in relation to interactions with skeletal elements, ligaments or muscles, as well as, in relation to specific functional/mechanical requirements. The only exception appears to be the teeth, which display a unique microstructural architecture composed of a magnesium-bearing calcite crystal combination, such as monocrystalline plate-elements, monocrystalline fibrous-elements and polycrystalline matrix, with a variable amount of organic macromolecules (about 0.2–0.25 wt %) [103, 118, 119, 204, 206–209]. Echinoid teeth are elongated, moderately curved and highly variable in shape, and can be classified in four types (U, T, prism and wedge-shaped teeth) on the basis of their different cross-sectional profile (figure 7(C)) [21, 173, 191, 205, 210]. Along the longitudinal axis, each tooth displays three main well-differentiated parts: an aboral growing portion (plumula), a midshaft and a mature portion characterized by a sharp oral tip [191]. In order to cope with the constant tip abrasion due to the interaction with the substratum, the tooth grows continuously at the plumula level and then slowly descends along the jaw following an inner pyramidal furrow [191, 211, 212]. The mature part consists of three main zones characterized by well differentiated structures and functions: (1) the primary plate zone, organized in lamellar plates and prisms obliquely oriented with respect to the longitudinal tooth axis; (2) the stone part, formed by calcareous needles surrounded by a polycrystalline matrix and connected to the primary plates by lamellae; (3) the keel, consisting mainly of inner prisms and of outer secondary plates with peculiar carinar prolongations [118–120, 131, 202–205, 213]. Echinoid teeth were analysed in detail using various techniques, such as SEM, energy-dispersive x-ray spectroscopy analysis, x-ray micro-tomography and spectromicroscopy, as well as micro- and nanoindentation, in order to identify their microstructure, material distribution, mechanical behaviour, and chemical composition. These analyses allow an interpretation of the tooth's structural architecture and integration in relation to its complex mechanical performance [118–120, 214]. In terms of structure–function correlation, the lamellar plate components appear to be a structural solution adapted to reinforce the zones subjected to maximum compressive stress (abaxial part), whereas the fibrous elements are employed in the zones of maximum tensile stress (adaxial part: the keel) [52, 120, 202, 215]. At the tooth tip, plates and fibrous elements split off due to shearing forces consequently creating a fracture at the surrounding organic layer, generating a mechanism for self-sharpening [119, 215]. Recently, this mechanism has been further investigated using 3D techniques in-situ SEM experiments and mechanical measurements combined with a nonlinear finite-element analysis [216].
In conclusion, the tooth is adapted to minimize and respond to multiple and combined mechanical stresses such as shear, bending, torsion and buckling produced by gripping, scraping, digging and locomotion [52, 119, 120, 204, 210, 215]. The strategic employment of magnesium-calcitic material together with its mechanical properties, in combination with the orientation of a plate-and-prism arrangement (according to the lines of force of the applied loads), result in a remarkable increase in tooth hardness (twice that of inorganic calcite itself) allowing echinoids to dig efficiently and deeply into calcareous rocks [93, 120, 201, 214, 215, 217–222].
5. Accessory appendages
Echinoids possess a variety of articulated accessory appendages [18, 32, 77] including spines, pedicellariae and sphaeridia. Spines and pedicellariae are primarily involved in defence and cleaning and can often show signs of damage and repair or can even be autotomised [223–225]. Sphaeridia are minute skeletal spheres attached to the test around the peristomial ambulacral regions (lacking in cidaroids) and are considered to be statoreceptor and proprioceptor organs. However, little is known about their morphology and physiology [39, 226, 227].
Pedicellariae are minute pincer-like structures distributed on the test surface, particularly around the peristome (figure 8) and periproct [34, 228] and are employed in different activities such as gripping, defence, covering and cleaning [34, 39, 47, 77, 229–236]. As most musculo-skeletal organs, each pedicellaria consists of a stalk, neck and two to five valves. Pedicellariae are highly variable in shape, often denticulated and sometimes armed with venom glands [77, 233, 237, 238]. Due to their variable shape, pedicellariae have been extensively used in taxonomy [5–11, 34, 239–241]. The valves show specific stereom structures and are equipped with functionally distinct muscles (abductors, adductors and flexors) and collagenous ligaments [242], which contribute to its gripping force [231, 233]. They generally react to chemical and tactile stimuli, in fact most valves are equipped with fields of chemosensitive cells [243–245]. As reported by Cavey and Markel [39], and further investigated by Coppard and co-workers [34], there are four main types of pedicellariae: (1) globiferous pedicellariae, which possess venom glands and denticulated valves with large and strong adductor muscles: they are employed as a deterrent against medium and larger predators; (2) ophicephalous pedicellariae, which possess three denticulated valves, provided with a glandular portion involved in releasing anti-fouling substances onto the test surface, and larger processes for muscle attachments enabling them to exert more strength and reduce muscle fatigue during object holding (figure 8(B)); (3) triphyllous pedicellariae, which are the smallest type of pedicellariae, are characterized by three small valves, long muscular neck and stalk: they are not sensitive to touch, have limited holding time and are employed to free test surface of minute particles (figure 8(C)); (4) tridentate pedicellariae, which are the largest and most common type, consist of three denticulated valves: they are activated by tactile stimulation and employed in removing larger particles or preventing test contamination by invertebrate pests. Past studies on pedicellariae generally consisted in descriptions of their morphology, activities and functions [228, 236, 246, 247]. Noteworthy are Campbell's studies that analyse in detail the forms and activities of the pedicellariae, identifying jaw movements, closing and opening responses (occurring after direct reflex-arc stimulation or indirect nerve stimulation), as well as, their latency, speed and duration, receptor distribution and reaction [229–233, 244].

Figure 8. SEM micrographs of pedicellariae. (A) Oral view of Paracentrotus lividus showing the peristomial membrane area including the mouth with five teeth, spines, tube feet and pedicellariae. Bar = 1 mm. (B) Detail of ophicephalous pedicellaria tip. Bar = 100 μm. (C) Detail of triphyllous pedicellaria tip. (B) and (C) are identified in (A) with dotted circles. Bar = 40 μm. p = pedicellariae, pm = peristomial membrane, t = tooth, tf = tube foot, sp = spine.
Download figure:
Standard image High-resolution imageSpines are elongated structures consisting of shaft (neck and tip), milled ring and base [80]. Each spine is joined to a respective tubercle at a ball-and-socket joint [80, 114, 248] and can be moved or firmly maintained in position due to the combined synergic action of a muscle and a ligament, known as the 'catch apparatus' [249]. The spine base enclosed by an articular envelope including a continuous outer layer of parallel muscle fibres runs from the spine to the test, and an inner layer of parallel ligament fibres with spine-test attachments. The ligaments consist of MCT [for review see 162, 250] enabled to drastically and quickly change its mechanical properties under nervous control. The presence of MCT allows the tensile state of the ligament to change rapidly from a soft and flexible condition, favouring muscle action during movement, to a rigid condition, locking the spine in position without muscle involvement, providing a fatigue/ energy-free holding mechanism [251]. Spine shape and size differ greatly from species to species: like a needle they can be long, hollow, thin and pointed as those in camarodonts; or look cylindrical or flattened, long or short, streaked or variously decorated, as in cidaroids; or moreover, appear modified and miniaturized as in irregular echinoids (figure 9) [29–33]. Spines perform different functions, such as locomotion, feeding and burrowing [29, 39, 42, 252]. They also act as a protection from physical trauma and predators [40, 253, 254] and as a stress impact reducer [43, 255–258], which is one of their main roles in the prevention of structural test damage. As reported by Tsafnat and co-workers [135] this is achieved by the spine microstructure, which improves resistance to compression. Thus, spines are structurally highly adapted to withstand different mechanical stresses, combining high impact resistance with high-energy absorption [43, 54, 59, 61, 65, 123, 132, 133, 135, 239, 258–261]. As for other skeletal components, the mechanical performance of the spines is the result of three hierarchical features, i.e. material properties, microarchitecture and shape. With regard to material properties, even if each spine behaves as a single calcite crystal with the c-axis oriented to its long axis [262]—as shown by polarized light microscopy [89], x-ray diffraction [107], and electron backscatter diffraction [132]—it has a mesocrystalline structure [99] consisting of a highly oriented array of nanocrystals embedded in a matrix of ACC and macromolecules [95–103, 110, 263–265]. The presence of ACC and intracrystalline macromolecules determines a typical conchoidal fracture behaviour resulting in increased fracture resistance and structural flexibility, as shown in the other skeletal parts [95–99, 110–113, 116, 121]. The material composition within the spine is highly variable (particularly the magnesium concentration), implying diverse mechanical properties in terms of elastic moduli, hardness and stiffness, and is significantly higher in the septa rather than in the spine central core [61, 132, 133]. Apropos the microarchitecture, spine stereom types greatly vary from species to species and along the same spine [54, 132, 133]. This leads to a very specific structural behaviour regarding the stress pattern distribution and resistance, as shown by the mechanical tests, such as three-point bending [54, 132, 266] and bulk compression tests [61, 133, 258]. Spine growth lines have also been shown to possess a mechanical significance and their presence could enhance resistance to larger force values [54, 59, 61, 67, 123, 133, 267–269]. Spines can display a peculiar morphology (widely recurrent in nature e.g. feather shafts and plant stems) consisting mainly of a hollow cylindrical porous structures, well known for their efficient mechanical advantage related to high strength-to-weight ratio [270]. In addition, many spines are characterized by sets of radial elements such as wedges [54, 59, 123], barbs and bridges, optimizing stress distribution [54, 65, 123, 135], increasing bending stress resistance [259] and preventing fracture propagation [43, 65, 132, 135, 271, 272]. In particular, in Centrostephanus rodgersii, a detailed analysis of spine behaviour under compression, tension and torsion loads by means of micro-CT scan and FE-Analysis has led to the identification of stress concentration patterns within spines and their role as mechanical supports [135].
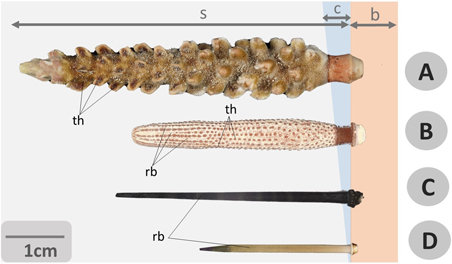
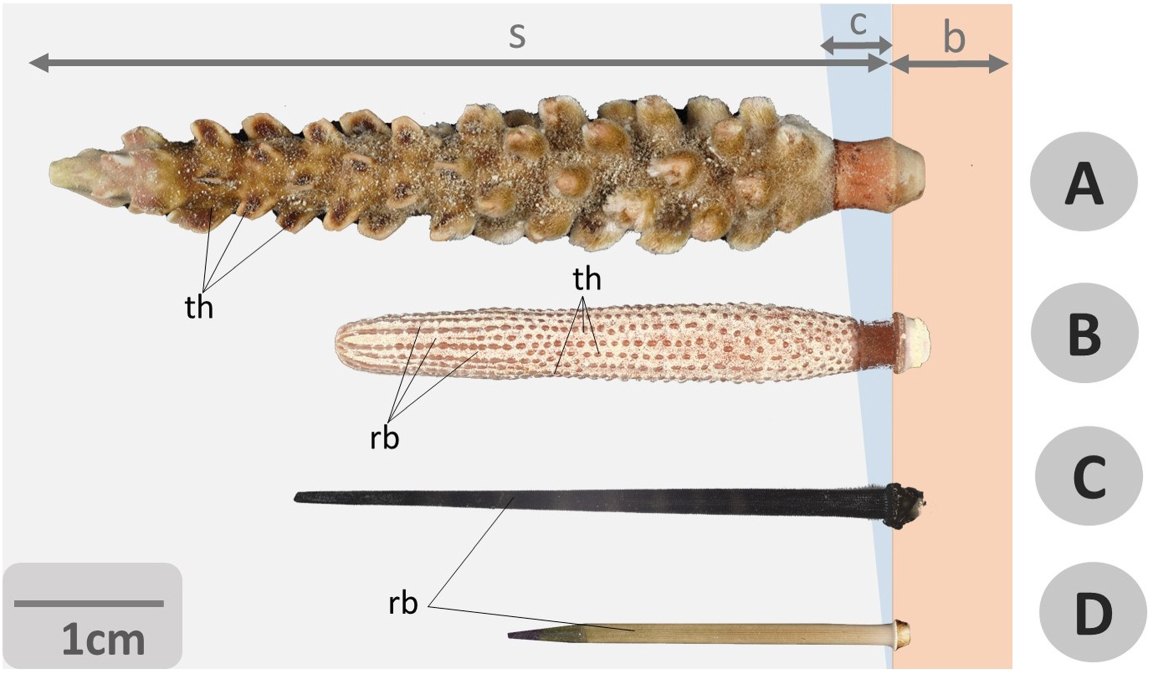
Figure 9. Photographs of echinoid spines. A general comparative view of spines from different species of cidaroid and camarodont regular sea urchins: relevant differences in shape, size and structure (including external, reliefs and tubercles) of shaft, tip and base are well evidenced. (A) Chandrocidaris giganteus (Cidaroida), (B) Phyllacantus imperialis (Cidaroida), (C) Echinothrix diadema (Camarodonta), (D) Echinometra lucunter (Camarodonta). Bar = 1 cm, s = c = collar, b = base, rb = ribs, s = shaft, th = thorns.
Download figure:
Standard image High-resolution imageIn addition to the true musculo-skeletal appendages, tube feet (or podia) may also be considered because they are intrinsically related to the endoskeleton and their soft wall can include skeletal spicules [273]. Each tube foot consists of an extensible hollow cylindrical stem with a terminal adhesive disc that includes small calcareous ossicles (rosette) [273–275]. Symmetrical pairs of podia (often used in taxonomic work) [276] emerge from the ambulacral plate test pores [25] and are moved hydraulically [277–280]. These podia are an integral part of the internal water vascular system whose major function is respiration [77], attachment [33, 277–282] and locomotion on different substrates [283], as well as, having a significant involvement in climbing, righting, covering [33], photoreception [284], food handling, burrow-building, and chemo/ mechano-sensory reception [285]. The specific tube foot attachment/detachment mechanisms are related to disc adhesive and de-adhesive secretions [281, 282], whereas stem extension, flexion and retraction are caused by typical hydraulic mechanisms [277–279] involving the employment of MCT [282]. As revealed by Santos and co-workers [280–285], the adhesive secretion is composed of inorganic material (45.5%), proteins (6.4%), neutral carbohydrates (1.2%) and lipids (2.5%). The adhesive strength (force per unit area) ranges from 0.09 to 0.54 MPa [277, 279, 282]. However, the adhesive and tensile strength and overall attachment capabilities change according to the number of tube feet involved [278, 279, 286, 287], species and population analysed [278], animal body size and shape, substratum and hydrodynamic conditions [255, 278, 279, 286, 287].
6. Biomimetic applications
The term biomimetics identifies an interdisciplinary approach that combines the understanding of natural structures, systems and processes with their abstraction and translation into technological applications [69, 71–74, 288]. Biomimetics is neither an imitation of nature nor a mere copy of forms, but rather it is an in-depth comprehension and translation of natural working-principles (e.g. constructional principles of organisms), which can optimize structures in building constructions, industrial products and technical processes [74–76].
The biomimetic process is supported by a series of analogies between biological and technical structures enabling the transfer of solutions on functional bases [289–292]. Indeed, organisms and artefacts are often faced with similar problems, such as the need to increase structural stability and resistance (skeleton/frame), pressure drag reduction (streamline shape and ribbing surfaces of marine animals/hull of boats) and reaction to external conditions (nastic movement of plants/dynamic facades) [45, 103, 293]. Hence, by understanding and modelling the adaptive principles of organisms, functional solutions for innovative design inspirations or 'bioinspirations' can be identified stimulating technical implementations [69, 71, 74–76, 289–292]. Nevertheless, the constructional design of organisms is subjected to different factors such as heritage constraints and morpho-functional adaptations to biotic and abiotic factors [28]. Hence, structural and functional solutions adopted by organisms are often neither the most advantageous nor the most adapted in any situation and context, since they represent a compromise respect to evolutionary constraints [28]. A specific contextualization and optimisation of biomimetic technical solutions is therefore required and can be performed through an interdisciplinary collaboration between biology and other scientific fields (e.g. engineering, design, architecture, material science, etc) with the aid of specific tools, such as 'computer-aided design' (CAD), 'computer-aided optimization', knowledge database and algorithms [294, 295]. Consequently, the abstracted, interpreted and contextualized biological principles can lead to new inspirations for the improvement of structures and/or processes based on analogies of functions [28, 69, 71, 74–76, 288–291, 295, 296].
On the other hand, biological structures significantly differ from artificial ones in various important aspects such as: growth process generating structures with full functionality and integrity at all stages of life [158, 297]; use of basic autochthonous and sustainable materials usually characterized by heterogeneity, anisotropy and hierarchy that determine multiple functions and emerging properties [45, 109, 158, 298]; integration in the environment and ability to interact with biotic and abiotic components [299]. On pair with analogies, these differences can also lead to new design perspectives and opportunities [66, 76, 288, 300], e.g. growing structures of material ecology [301, 302] responsive dynamic façades for building constructions [288], and hybrid design products [303, 304].
The biomimetic procedure is carried out in different steps and tools [for review see 74]. Although the methods adoptable in this field are different and numerous, they can be allocated in two types of approaches: bottom-up and top-down [305, 306]. The bottom-up approach begins by identifying adaptive functional solutions in biological species, followed by the identification of the most suitable design and technological area for their transfer. This approach in literature has also been defined in diverse ways: solution-based, solution-driven, biology push, biomimetics by induction and biology to design. The top-down approach begins from the analysis of complex technical problems to the pursuing in nature of biological models offering novel solutions. In literature, this approach is also also known as: problem-driven, problem-based, challenge to biology, technology pull and biomimetics by analogy [69, 71–74,305, 306].
A general bottom-up is here simplified in five key steps [74]; in addition, a case study on Paracentrotus lividus's test is used as an example [141, 165]. (1) Identification of a biological model, usually identifiable on the basis of an analogy of function and of problem solving related to the final application: P. lividus test model as a shell structure enduring environmental mechanical stress. (2) Investigation of the structure, performance and function of the biological model: FEA of the discontinuous shell structure divided into rigid hexagonal plates and semi-flexible sutures. (3) Abstraction of functional mechanisms and design principles: a discontinuous shell structure with semi-flexible joints reducing bending moments. (4) Modelling and analysis to determine and optimize the artificial system based on the biomimetic design principle: developed in design sketches and concepts, the abstracted principle is then implemented in a 3D model of a bioinspired shell structure. The technological implementation includes the choice of material, form and structure supported by virtual simulations and validations, with particular attention and adaptation to the final use. (5) Application and manufacturing of the bioinspired artefact: realization and final acceptance test of the pavilion inspired by P. lividus test (figure 10).

Figure 10. Biomimetic process. A five-step scheme of the biomimetic solution transfer: from a biological model to a technical application as exemplified by the Paracentrotus lividus case study. (1) Identification of a biological model: P. lividus test. (2) Investigation of structures, performances and functions of the biological model: a finite element analysis of the discontinuous shell structure divided into rigid hexagonal plates and semi-flexible joints. (3) Abstraction of functional mechanisms and design principles: a discontinuous shell structure with semi-flexible joints reduce bending moments. (4) Modelling and analysis determining and optimising the artificial system based on the biomimetic design principle: a 3D model of a bioinspired plexiglass open shell. (5) Biomimetic design application: the final implementation, manufacturing and testing of a bioinspired building construction (images designed by Gabriele Pontillo, Simona Landi, Lisa Vitale, Ilenia Rubino, Pasquale Ciardiello, Alessandro Fusaro of University of Campania 'Luigi Vanvitelli').
Download figure:
Standard image High-resolution imageThis process is not frequently linear due to constraints, context and scaling difficulties [288]. In this regard, the dimensional scale is a crucial factor: organisms have highly different working principles based on their dimensional realm [109, 307]. A direct scaling of the biological solution to the design dimension is not always possible, particularly in building constructions that concern not only size but also materials, external loading, life cycle, required safety range etc [288]. For this reason, the abstracted principles need to be usually translated, redesigned and contextualized to be successfully applied as new technical solutions [74, 305].
In all these approaches, knowledge integration and interdisciplinary methods and tools are essential for investigation and designing of biologically inspired structures. The study of biomimetics embraces both life and engineering disciplines [72, 289, 308]. Although, the functional characteristics and processes of nature conducting to the design of new innovative artefacts are immeasurable (e.g. bio-mineralization, growth processes and regeneration), bio-mechanical aspects are the most studied and implemented in the biomimetic field. A series of mechanical principles based on physical-mathematical laws appear to govern the structure–function relationship in organisms, as in artificial structures [45, 288]. Hence, the physical-mathematical approach can successfully describe bio-structures and their mechanical problems and performances. As shown by d'Arcy Thompson (1917), this biomechanical approach has been applied for decades [109, 158, 292, 293, 309–311]. Nevertheless, the contemporary advances in computational imaging acquisition, virtual simulation and manufacturing, together with the increased instrumental biological analysis resolution, lead to new developments for inter-disciplinary mechanical studies and biomimetics [71, 164, 295, 312]. Both biological structures and principles can be digitally analysed in depth at a micro- and nanoscale and better transferred into a multitude and various constructions and industrial products [295, 301–304, 313]. Consequently, biological structures are converted and analysed as 2D/3D models and directly connected to the technical process, becoming archetypes and/or guides for the genesis of the products [66, 295]. This creates a supporting process with efficacious tools for designers, engineers and scientists in the transition from real (organism) to digital (2D/3D archetype) and from digital (3D model) to the real entity (physical building, device or product), involving digital manufacturing techniques, which reproduce in a rigorous and functional way the analogous strategies and mathematical laws of nature [289].
These biomimetic methods and tools enable not only a successful transfer and unique application, but also a deeper understanding of biological structures, their bauplan and evolutionary process. This enhancing knowledge of the biological realm based on biomimetic approaches is referred to as 'reverse biomimetics'. In particular, it can be conceived as an interactive spiral where the results achieved by the biomimetic approach lead to a more detailed understanding of the biological systems, representing the basis for further investigation and conducting to eventual new transfers and developments in biomimetic products [305].
In this complex framework, the skeletal components and mechanical properties of the echinoid constructional design have revealed a high potential in transferring functional bioinspired solutions into new diverse technical applications [27, 55–58, 64, 258]. Recent studies have shown how the echinoid structure can be digitally investigated generating 3D models and applying FE-analyses to identify possible structural and mechanical principles [54–58, 64, 138, 216]. In addition, based on their primary function, skeletal components have found a major and coherent field of technological application from engineering and architecture to robotics, biomedical and material sciences.
6.1. Engineering and architecture
Echinoids have a long history as inspiring models for engineering structures. This interest has recently increased, in particular regarding rotationally symmetrical constructions, defined as echinodomes [314, 315]. Detailed analyses of these structures including their mechanical advantages and limits have been technically described and generally well understood. Different load conditions, such as self-weight, snow loads, wind and hydrostatic loads, which can generate over- or under-pressure, can be calculated adapting constructions to specific mechanical needs and functions [315]. Echinodomes have been applied to several constructions including long-term storage containers for gas and liquid fuels such as automobile and aircraft gasoline, mineral oil, and other volatile substances [315]. The advantages of echinodomes are specifically due to their thin-shelled and double-curved architecture that results in mechanical behaviour predominantly following the membrane theory, i.e. in-plane membrane stress, reduced bending stress [315–318].
Additional studies have not only focussed on the overall shape of an echinoid test, but also on specific working principles that have recently been implemented in civil engineering. Grun et al [64, 319, 320] provided an overview on echinoid skeletal strategies in building constructions, by identifying in the skeleton various structural working principles on different hierarchical levels and their transfer into demonstrators. These are architectural constructions providing a proof-of-concept of specific functional aspects. Transferred structural principles based on echinoid skeleton include: (1) mosaic-arranged plates, where three plates meet in one point in order to avoid straight edges, which may cause kinking; (2) clypeasteroid-type plates, interconnected by skeletal protrusions leading to secure plate interlocking; (3) fibre-connected plates; (4) light-weight constructions; and (5) double-wall constructions as found in Clypeaster rosaceus [27, 64].
Both structural elements and processes leading to specific echinoid morphologies have been investigated [64]. Plate distribution has been optimized using the echinoid skeleton as a role model [64, 321] and high-performance structures, identified and analysed, have been abstracted and transferred in various ways into demonstrators. For example, the ICD/ITKE Research Pavilion 2011 (figure 11(A)) [64, 321, 322] has well demonstrated the application of three structural principles among those cited above: (1) mosaic-arranged modules, where three modules meet in one point; (2) single hollow modules, made from multi-elements reflecting a lightweight construction; (3) modules interconnected by comb-joints. Similarly, a building construction in the form of the Landesgartenschau Exhibition Hall 2014 was realized (figure 11(B)) [64, 321, 323–326]. A second ICD/ITKE Research Pavilion developed in 2015 (figure 11(C)) focussed on (1) modules arrangement; (2) comb-joint refinement; (3) material differentiation using textile connections; (4) light-weight construction; (5) a double-shelled structure; (6) an evolutionarily optimized growth algorithm based on the echinoid growth process by plate addition [64, 321]. In 2018, the Rosenstein Timber Pavilion was exhibited demonstrating further developed, high-performance characteristics based on echinoid skeleton, focussing on improved plate connections and optimized plate distribution [323]. Furthermore, these characteristics have also inspired the BUGA Wood Pavilion (2019, ICD/ITKE University of Stuttgart) (figure 11(D)), which was realized combining a new digital design approach for shape-funding structures with an automated robotic manufacturing using wood, thus receiving the German Design Award 2020 in the 'Excellent Architecture' category [327]. As a final example, the Rosenstein Pavilion was realized in 2019 as a functional graded concrete shell structure inspired by the stereom of Heterocentrotus mammillatus spines. In particular, the spine structure was investigated as a main biological model for the designing of a new functional graded porosity of a concrete shell. The abstracted principle lead to a structural efficiency improvement of the porous pavilion through a functional distribution of material in accordance to a dominant stress state, resulting 40% lighter [328].

Figure 11. Echinoids in Architecture. Research pavilions and building constructions inspired by both morphological and mechanical principles of the genera Clypeaster and Phyllacantus. (A) ICD/ITKE Research Pavilion 2011. (B) Landesgartenschau Exhibition Hall 2014. (C) ICD/ITKE Research Pavilion 2015–16. (D) BUGA Wood Pavilion 2019 (© Photo A–D curtesy of Prof. Jan Knipper, curtesy of Prof. Jan Knipper, Institute of Building Structures and Structural Design, University of Stuttgart.
Download figure:
Standard image High-resolution image6.2. Robotics
Various studies were carried out in the robotic sector from the analysis of echinoid biology and structures to the development of new robotic designs [329]. As an example, a sea urchin-like robot was designed as a new exploration platform enhancing access to unstructured environments or dangerous places [330]. Based on tube feet and spine locomotion a flexible spherical rolling robot was developed with retractable linear actuators and pendulum-driven mechanisms. Both strategies intended to overcome the locomotion difficulties of spherical robots on irregular surfaces [330]. Echinoderm tube feet have been a source of inspiration for a wide range of soft robotic actuators [331–333]. For example, studies based on tube feet models have resulted in a magnetically controlled crawling mechanism [334] and suction device optimized for grasping rough surfaces with a rapid release mechanism [335].
An interdisciplinary team of engineers and marine biologists from the Jacobs School of Engineering (University of San Diego, California USA) used the Aristotle's lantern to develop a space exploration robot with a new gripping device for sediment sample collection (figure 12) [336]. Starting from the analysis of the opening and closing mechanism of the lantern system and the bio-exploration of keeled and non-keeled teeth, a bioinspired model was built and tested via FEA determining the efficiency of the lantern-like mechanism and confirming the structural importance of the keel in the reinforcement of the sea urchin's tooth [336].

Figure 12. Echinoids in robotics. Gripping device for sediment sample collection inspired by Aristotle's lantern. Computer aided design images of the bioinspired device (A) closed, (B) fully opened. (C) 3D printed bioinspired device in collecting different types of sand on the beach. This image is adapted from [336].
Download figure:
Standard image High-resolution image6.3. Biomedical engineering
An optomechanical biopsy device for minimally invasive surgery was realized [337] adopting the lantern's ability to simultaneously scrape and engulf food in alternating and combined movements of opening/protrusion and closing/retraction following Scarpa's pioneering bionic model [338, 339]. The prototype was implemented as an extrudable steel tube (0.15 mm thickness and 4.3 mm diameter) provided with a cutting device, i.e. a crown-shaped system characterized by triangular teeth, designed to perform an accurate biopsy in less than a millisecond (figure 13) [337].
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 13. Echinoids in bioengineering. Optomechanical biopsy device inspired by Aristotle's lantern. (A) Drawing of the Aristotle's lantern bionic model. (B) Crown-shaped system characterized by triangular teeth in closed and opened positions. (C) Prototype implemented as an extrudable steel tube thick 0.15 mm and 4.3 mm in diameter. (D) Drawing of the bionspired device during surgical activity. This image is adapted from [336].
Download figure:
Standard image High-resolution image{kind=link}
In the biomimetic industrial design field, especially in the biomedical sector, a recent study on the mechanical design of P. lividus test was carried out by an Italian team (Hybrid Design Lab, University of Campania 'Luigi Vanvitelli' and Department of Structures for Engineering and Architecture, University of Naples Federico II) [141, 165, 340]. As an example, the identified adaptive solutions of the test, as a modular system guaranteeing high integrity and structural stability in different stress conditions, were transferred into the design of two different biomedical devices: an arm-tutor and a cranial harmonizer. Shape and structure of the biological models were abstracted and applied, according to principles of functional analogy, and reproduced in parametric 3D CAD models responding to specific innovation needs expressed by users and medical experts, these are: (1) lightness, ensured by a controlled porous arrangement mimicking stereom structure; (2) resistance and stability, obtained by a discontinuous structure consisting of hexagonal modules connected by semi-flexible material reflecting the modular plated structure of P. lividus test and its low flexural stiffness at the sutures; (3) breathability, ensured by the high structural porosity and modular subdivision, reducing the presence of closed spaces; (4) free customization for different therapeutic needs and personal preferences, provided by an elevated versatility of shapes, geometries, colours and styles obtainable by parametric designs and digital manufacturing [340].
Pedicellariae-like devices have also been developed into new versatile tools in micromanipulation and micro-robotics fields for healthcare. Leigh and co-workers [341] designed bioinspired forceps using micro-stereolithography creating a pneumatic chamber that opens and closes the jaws by changing pressure using a syringe. The device can be used for functional grasping of microparticles and in addition can be activated hydraulically exhibiting a self-healing behaviour (isolating the damaged regions and maintaining the hydraulic mechanism efficiency) [341].
6.4. Material science
Echinoid spines revealed an important potential for innovative bio-inspired applications due to their sophisticated lightweight structure and material properties, in combination with strategic failure behaviour, high impact resistance and high-energy absorption [59, 61, 65, 133, 258, 342].
In particular, the calcitic microstructure of echinoid spines inspired: (1) new porous biomaterials useful for prosthetic applications, especially for synthetic teeth and bone replacement [343]; (2) optimized microporous vascular prostheses [344]; (3) bone implant materials obtained from the conversion of sea urchin spines to Mg-substituted tricalcium phosphate [345]; (4) model preparation of new materials by combination of crystals, polymers and functional molecules [135]; (5) a macroporous copper with greater mechanical properties [346]; (6) new functionally graded concrete [258].
In particular, the echinoid microstructure was deeply studied as a functional model to create new prosthetic materials. During the 70's, Weber et al [343] successfully replicated the skeletal structure of the Heterocentrotus spines in epoxy resin and in sodium silicate. In particular, they recognized in the arrangement of the echinoid 3D microstructure some important characteristics, which transferred into new functional prosthetic materials, were able to provide structural strength and proficient surface for tissue growth. In this regard, the stereom was identified as an optimized construction ensuring a good permeability and functional porosity, as well as a periodic minimal surface structure, in which the interface between calcite and the organic phase offers maximum contact for crystal growth [116]. Following studies involved a direct conversion from echinoderm material to bio-implant materials [345, 347]. In particular, based on a hydrothermal conversion, the spines of the echinoids H. mammillatus and Heterocentrotus trigonarius have been converted in Mg-substituted tricalcium phosphate for bone implant, maintaining the interconnected porous structure with a good bioactivity and osteoconductivity. Currently, high-resolution and advanced techniques in tissue engineering are able to reproduce new artificial scaffolds with a controlled porosity at micro and nanoscale; thus, these bioinspired solutions can be more effectively transferred creating new opportunities to realize innovative synthetic or hybrid materials [348, 349].
In addition, different studies on the cidaroid Phyllacanthus imperialis and H. mammillatus spines were carried out, showing how the specific arrangement of porous material, associated with different densities and architectures, allows these species to have extremely light and resistant structures identified as ideal models for the realization of new aluminium ceramic and concrete materials [59, 65, 67, 139, 261].
Lightweight structural ceramics have also been developed using the echinoid skeletal plates to template the synthesis of effective porous materials. As an example, porous gold structures with nearly regular 15 μm channels were prepared by coating skeletal plates with gold and dissolving them and leaving the original structural form [350, 351]. These materials with a pore dimension comparable to optical wavelengths could be applied for their optical properties or used as catalyst supports.
These examples highlight how biological principles can be successfully abstracted and transferred into technical applications [308]. Moreover, in a reverse biomimetic view, these analyses also provided a more detailed insight on morphology, function and integration of an organism in its ecosystem [46, 58, 269]. In particular, this allows a better understanding of an organism's adaptation to its environment, the evolutionary pathway of its structure, and its ecological and paleontological implementation [352, 353]. For example, the comprehension of the structural design, skeletal strength and weaknesses of the echinoid test consents to interpret taphonomic processes and the potential preservation of the echinoid taxa [56]. Such knowledge can help ecologists and palaeontologists to better assess the effect of taphonomic filters and biases on echinoid communities helping to determine e.g. if predatory drill holes or other biotic traces can promote the potential preservation of an echinoid [354] or lead to a loss of information.
7. Conclusion
In the course of time, the original constructional design of the echinoid endoskeleton has attracted the attention of researchers from different scientific fields due to its unique morphology, structure and material properties. Currently, these features reveal a great potential for biomimetic applications, thus motivating further investigations. This review presents a comprehensive synthesis of important studies on mechanical design and principles of echinoid skeletal structures, emphasised the efficiency of the endoskeleton at different hierarchical levels. Each constructional element of the echinoid's skeleton demonstrated to have a major application as a biological role model: the test in building construction; Aristotle's lantern and pedicellariae in grabbing devices; tube feet in robotic locomotion systems; spine stereom and biomineral composition in innovative materials. Contemporary technological advances in computational imaging, numerical simulation and fabrication have paved the way to a new era for the study of mechanical principles in organisms and their functional transfer [64, 295, 301–304, 340, 355]. Mechanical strategies and performances of the various components can be highlighted by means of different types of digital advanced techniques, such as high-resolution x-ray microcomputed tomography, imagine analysis, 3D modelling and FEA. These technologies ensure high fidelity in the acquisition of biological models, great reliability of results and high reproducibility of complex geometry and structures through the new frontiers of digital manufacturing techniques [64, 164, 301–304, 319, 356, 357].
Consequently, a new virtual biology is emerging capable to provide novel answers to questions concerning the morphology, function and evolution of living and fossil species [164, 356, 357]. In this regard, studies of mechanical design in organisms are just at an initial phase. Nonetheless, according to present literature, there is evidence of a significant increase in research [46, 55, 57, 59, 124, 135, 217, 218] regarding new future integrations between cutting-edge computer science and biology. In conclusion, this review aims to illustrate how the constructional design of echinoids reflects animal adaptations to specific mechanical needs related to different environmental stresses and lifestyles, which abstracted and transferred into engineering and industrial design, provide functional solutions improving structures, processes, and human health.
Funding
This work was supported by the Department of Engineering, University of Campania "Luigi Vanvitelli", PhD program [National Operational Program (PON) "Research and Innovation" (R&I) 2014–2020]. Tobias B Grun's work was supported by National Science Foundation grants EAR SGP-1630475 and EAR SGP-1630276.
Acknowledgments
The Authors thank Prof. Arch. Mario Buono (University of Campania Luigi Vanvitelli, Aversa, Italy), the Zoological Station Antorn Dohrn (Naples, Italy) and the Hybrid Design Lab, (Naples, Italy) for their support. They also express thanks to John Slapcinsky (University of Florida, Florida Museum, Invertebrate Zoology) for the access to the echinoid spine collection; Ms. Laurajean Carbonaro-Tota for the English revision.