

Abstract
Studies assessing potential measures to counteract the marine deoxygenation attributed to anthropogenic activities have been conducted in a few coastal environments and at regional scale, but not yet on a global scale. One way toward global scale artificial oxygenation would be to use oxygen produced as a by-product from hydrogen-production through electrolysis. The low-carbon footprint renewable production of hydrogen from offshore wind energy offers such a possibility. Here, we assessed the potential of this artificial oxygenation method on a global scale using a coupled physical-biogeochemical numerical model. The anthropogenic oxygen source scenario assumes worldwide adoption of hydrogen, considering demographic changes and the feasibility of offshore wind turbine deployment. Following this scenario, artificial oxygenation had a negligible effect on the overall oxygen inventory (an increase of 0.07%) but showed a reduction in the overall volume of Oxygen Minimum Zones (OMZs) between 1.1% and 2.4%. Despite the decrease in the mean OMZ volume globally, OMZs display distinct and contrasting regional patterns notably due to the oxygen impacts on the nitrogen cycle. Artificial oxygenation can inhibit denitrification resulting in a net gain of nitrate that promotes locally and remotely increased biological productivity and consequent respiration. Increased respiration could ultimately lead to an oxygen loss at and beyond injection sites as in the Tropical Pacific and Indian Ocean and particularly expand the Bay of Bengal OMZ. In contrast, the tropical OMZ shrinkage in the Atlantic Ocean is attributed to oxygen enrichment induced by advective transport into the OMZ, while the absence of denitrification in this area precludes any biochemical feedback effect on oxygen levels. These results suggest that the impacts of artificial oxygenation on oxygen concentrations and ecosystems are highly non-linear. It can produce unexpected regional responses that can occur beyond the injection sites which make them difficult to forecast.

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Oxygen availability is important to marine ecosystems notably because of its control on biological respiration and organic matter (OM) remineralization. The dissolved oxygen (O2) supply to the ocean occurs through atmospheric air–sea gas exchange and release from photosynthesis [1]. While oxygen is abundant in the upper ocean, vast zones of the ocean display naturally low oxygen concentrations known as Oxygen Minimum Zones (OMZs). These OMZs occur in poorly ventilated areas, below productive surface regions (between 200 and 600 m) and are defined by O2 thresholds representing different stress levels on the marine ecosystem. In this study, we followed Bopp's and Cocco's definition [2, 3] with thresholds defined as , and , corresponding to distinctive effects on biogeochemistry (respectively oxygen stress for marine organisms including Atlantic Ocean's species, hypoxia: a shortage of oxygen for life processes, and suboxia: O2 conditions conducting to denitrification and anaerobic ammonium oxidation). OMZs have been shown to expand in the context of a global ocean deoxygenation attributed to global changes [4–8]. The upper ocean has lost 0.5%–3.3% of oxygen since the 1970s [9]. Moreover, the deoxygenation and OMZs expansion is expected to continue throughout the 21st century. This additional deoxygenation taking place over the next 100 years is estimated between 1%–7% of the current global inventory according to ocean climate model projections and different climatic scenarios [2, 10], but is potentially underestimated according to the recent Coupled Model Intercomparison Project (CMIP6) [11]. In addition, decreasing marine O2 has been shown to intensify marine hypoxia, thereby compressing the habitat of aerobic organisms [12], influencing biogeochemical processes (e.g. the remineralization of OM and the nitrogen cycle, [13]) and ecosystem diversity, leading to potentially severe socioeconomic impacts [14].
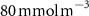
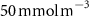
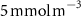
Although the ultimate solution would be to drastically reduce anthropic greenhouse gases (GHG) and nutrient pollution, anthropogenic reoxygenation has been proposed as a potential mean to mitigate deoxygenation. Despite the uncertainties regarding the origin of hypoxia, engineering methods have been tested to mitigate their expansion (with modeling and in-situ approaches) in some lakes [15] and coastal marine environments, with contradictory results [16]. For example, similar conceptual oxygenation techniques (artificial downwelling of oxygen-rich water) were carried out in two different regions resulting in either an in situ deep water oxygen restoration in the By Fjord [17] or a modeled Pacific desoxygenation and changes in biological activity beyond the experimental sites [1].
An evaluation of the direct O2 injection effects at global scale has, to our knowledge, never been carried out. Thanks to the electrolysis of water used to produce green hydrogen, which generates O2 as a byproduct, the injection of O2 is now conceivable at global scale into the ocean. Indeed, hydrogen produced by water electrolysis from renewable energy (e.g. from offshore wind farms) is now widely seen as green hydrogen given its low environmental footprint [18] and has therefore been recently subjected to high investment by developed countries and foreseen to be widely used among the global population.
The current study aims to assess the potential of this anthropogenic oxygen (AO) injection into the ocean and its environmental impact using a specifically designed global scenario with a coupled physical-biogeochemical model.
2. Experimental setup
2.1. Numerical models used
We used the Nucleus for European Modelling of the Ocean (NEMO) platform (version 4.0). The NEMO platform comprises an ocean dynamical (NEMO-OCE in a ORCA2 configuration [19]), a sea ice (NEMO-ICE) and a biogeochemical model (NEMO-PISCES) to simulate the ocean at global scale. The biogeochemical component, named Pelagic Interaction Scheme for Carbon and Ecosystem Studies (PISCES-v2), is an intermediate complexity marine ecosystem model simulating the main biogeochemical cycles and the first trophic levels. Four main living compartments are simulated (two phytoplankton and two zooplankton functional groups) as well as detritus (including two size classes, small and large organic particulate carbon (POC)), nutrients and O2 [20]. As an extensive description and validation of the model has been published in [20], therefore only the relevant parts of the model are recalled here.
In the model, air–sea O2 exchange is computed using the commonly used Wanninkhof's parameterization [21]. O2 concentration in the ocean depends mostly on photosynthesis (O2 production) and remineralization (O2 consumption) with a O2/C Redfield ratio equal to 1.34 [22]. Denitrification process (i.e. the use of the oxygen from nitrate (NO3) when O2 concentration lower than 6 µM [23]) is explicitly simulated in PISCES.
The relative complexity of the O2 cycle in the NEMO-PISCES model results in good abilities to (i) represent volumes of low oxygenated waters at global scale [2] (figure C1(b)), and (ii) simulate a reasonable mean state compared to other global biogeochemical models [24, 25] used in CMIP. These good performances explain our choice of using the NEMO-PISCES model for the present study.
2.2. Simulation setup
We used the PISCES model forced offline by an ocean dynamical seasonal climatology as in [20]. The model spatial horizontal resolution is 2∘ by 2∘ cosφ (where φ is the latitude) with a meridional resolution refined to 0.5∘ near the equator. The vertical grid is composed of 31 layers of widths varying between 10 m near the surface to 500 m at depth. Prior to our experiments, we have spun up the biogeochemical model for 2000 years, starting from observed climatologies for nutrients [26] and from uniform low values for ecological tracers, until quasi-equilibrium was reached. After this spin-up, 100 additional years were simulated in two different configurations: (1) a control simulation (CTRL) without AO perturbation, (2) a simulation with an added AO (OXYBIO). A third configuration, named OXYNOBIO, simulates AO as a conservative passive tracer regardless of the oxygen cycle, with the exact same source of AO than the one used in OXYBIO. The aim of simulating the OXYNOBIO tracer is to track the transport of AO over a span of 100 years.
The experimental scenario used for the AO injection was based on the theoretical economical viability and industrial feasibility of offshore implantation of hydrogen Production Units (PUs). Offshore locations have been gauged considering socioeconomic and technological data mainly in line with the forecasts from the International Energy Agency [27]. These theoretical locations were provided by the LHYFE company [28] which is a producer and supplier of green hydrogen. This idealized scenario was designed to give an upper bound of an AO injection by assuming a global adoption of hydrogen as a key component of renewable energy supply chains by 2030, which corresponds to the highest level of plausible green hydrogen use.
For this study, the assumed number of hydrogen plants installed over the next 50 years follow roughly the same deployment than the offshore oil platforms between 1975 and 2020: approximately 20 000 platforms. A scenario allowing the deployment of 17 936 PUs has therefore been defined. The PUs have been distributed within 304 selected model grid points (each covering about 400 000 , called hereafter 'stations'), resulting in an average of 59 PUs per model grid point. These stations were distributed along the world's coasts (figure 1) according to the combination of two criteria: (i) an average wind speed preferably between 12–17 (optimal operation of the wind turbines) or between the minimum and maximum wind speed needed to operate the wind turbine (5–25 [27, 29]), and (ii) a population density that is presumed to exceed 2000 people (related to the size of the energy market), or display a constant or even positive natural evolution of the population by 2050 [27, 30]. As a result, the locations of the PUs are not necessarily positioned near known OMZs. Finally, we chose an injection depth of 150 m, except in areas with shallower bathymetry where the AO was injected at the greatest depth possible. Indeed, injection at a depth exceeding 150 m (e.g. close to the core of the OMZ) would have been more costly and would therefore not be economically viable. Nevertheless, the depth chosen here remains close to the upper limit of the OMZ [31] (figure C1).
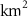
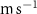
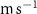
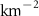

Figure 1. Spatial distribution of (a) annual anthropogenic oxygen flow injected by each station (see section 2.2) and (b) anthropogenic oxygen stations (orange dots) with the 'correspondence' (R) between and in background, where R∼0 : the dominance of AO dynamic transport over biological activity, R∼1 : the transport and the biogeochemical processes are of similar importance, R>1 : the dominance of biological activity over dynamic transport (for more details on the method, see appendix
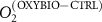
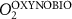
Download figure:
Standard image High-resolution imageCurrently, the green hydrogen production can reach or even exceed 129 000 t H2 , depending on the PU [28]. During the production of hydrogen by electrolysis, the H2/O2 ratio is 2:1 [32]. This amounts to 8 kg of O2 per 1 kg of H2 produced. Thus, according to LHYFE's forecasts, each of the PUs would produce 70 000 t O2 (figure 1(a)), i.e. 39.2 globally, which corresponds to 0.86% of the overall biogenic O2 production in the CTRL simulation between 0–150 m (4.56 ).
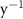
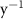
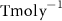
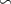
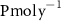
It is important to note that the idealized setup of our experimental design leaves aside engineering injection details (discussed in section 3.5) that may impact the efficiency of the AO injection.
3. Results and discussion
3.1. (De)oxygenation and OMZs widespread changes induced by anthropogenic oxygen injection
After 100 years of AO injection (OXYBIO run), the global O2 inventory increased by 0.07% with respect to the CTRL run (estimated to be 233.1 Pmol, a value close to that obtained from the observations by [8]: 227.4 Pmol). Yet, the AO injection reduced the volume of OMZs by between 1.1% and 2.4%, depending on the O2 concentration threshold used (table 1). This suggests that the overall evolution of the OMZ volume is weakly sensitive to the chosen threshold. However, the global average hides significant regional disparities.
Table 1. The proportion of the volume of each OMZ to the overall volume of OMZs (volume contribution) at the last simulated year of the CTRL run and averaged between 200 and 600 m depth, and change (OXYBIO minus CTRL) in volume of the oxygen-poor water (generically referred to as OMZs) after 100 years of AO injection. The volumes are defined by oxygen thresholds , and . Free fields (*) represent the absence of water masses with O2 concentrations below and bold indicates values for OMZs at global scale (sum of OMZs).
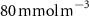
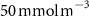

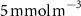
| OMZ | ||||||
|---|---|---|---|---|---|---|
| % | Change | Volume contribution | Change | Volume contribution | Change | Volume contribution |
| Global | −2,4 | 100 | −1,1 | 100 | −1,1 | 100 |
| Atlantic Ocean | −13,3 | 6,6 | −34,6 | 4 | 0 | 2,4 |
| Pacific Ocean | −2,3 | 81,3 | −0,6 | 86,1 | −2 | 94,6 |
| Bay of Bengal | +5,5 | 8,1 | +11,6 | 7,9 | +25,3 | 3 |
| Arabian Sea | −3,4 | 4,1 | −6,1 | 2 | * | * |
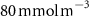
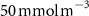
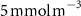
The OMZs that shrank the most are the least extended (figure 2(a), table 1): in the Atlantic Ocean and the Arabian Sea (AS), that shrank by 13.3%–35.6% and 3.4%–6.1% respectively ( and thresholds). The smallest shrinkage (0.6%–2.3%) is that of the Pacific Ocean OMZ (table 1). The most striking consequence of the AO injection is simulated in the Bay of Bengal (BoB) where the OMZ expands counterintuitively, its volume increases by 5.5%–11.6% for the and thresholds, and even 25.3% for the threshold (table 1 and figure 2(b)).
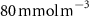
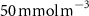
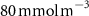
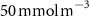
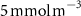

Figure 2. (a) Spatial distribution of the annual mean O2 concentration of the last simulated year of the CTRL run (mmol m−3) averaged between 200 and 600 m depth, the typical depth range of the main OMZs. The area of concentration below is outlined in grey. (b)–(f) Changes in annual mean O2 concentration (OXYBIO minus CTRL) after 100 years of AO injection (b) averaged between 200 and 600 m depth (), and (c)–(f) along meridional sections (%), in (c) the Atlantic Ocean at 2 ∘E, (d) the Pacific Ocean at 120 ∘W, (e) the Bay of Bengal at 90∘E and (f) the Arabian Sea at 70 ∘E. In (c)–(f), the isopleths at 50 and are shown in blue for the CTRL simulation and in orange for the OXYBIO simulation, and the black solid lines show the isopycnals that delimit the water masses based on water masses definition in the appendix
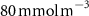
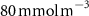
Download figure:
Standard image High-resolution imageContrary to the global assessment, the volume variation of regional OMZs depends strongly on the threshold that is considered to define them. The harshest thresholds specific to each region allow us to determine the evolution of the OMZ core. Thus, excluding the Pacific OMZ, the volume change is the most relevant in the OMZ core (i.e. a larger volume variation for the threshold of in the Atlantic Ocean and in the AS and BoB than for , table 1). However, the evaluation of the structure of an OMZ requires to also consider its upper and lower boundaries (oxyclines) [31]. OXYBIO configuration simulated a slight deepening of the upper oxycline of all OMZs (figures 2(c)–(f)), more marked in the Atlantic Ocean. Thus, their thickness thins by an average of 42 m (for the Atlantic Ocean) and 13 m (for the AS) between 0 and 600 m while that of the BoB thickens by 11 m despite the substantial increase in O2 between 100 and 200 m, which can exceed 100% (figure 2(e)). The shift of the horizontal boundaries position of the OMZs in addition to the deepening of the vertical boundaries, characterize the strongest shrinkage of the OMZs, namely in the Atlantic Ocean, and to a lesser extent in the AS (figures 2(c) and (f)).
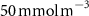
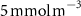
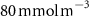
3.2. Mechanisms involved in the evolution of OMZs
A wide range of ecosystem processes interacting with the three-dimensional circulation determine the distribution of O2 concentrations in the ocean [33]. The analyses in section 3.1 suggest a regional disparity in the mechanisms causing O2 variation. (AO in the OXYNOBIO configuration) tracks exclusively the transport of AO without any air–sea exchanges and biogeochemical reactions. The correspondence (R) between the variation of and that of (i.e. O2 change) addresses the relative importance of biological activity and transport to O2 variation in OXYBIO (figure 1 (b), equation (1) and appendix
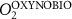
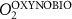
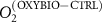
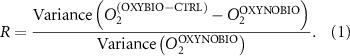
R contrasts O2 changes due to biological response (i.e. minus ) to AO transport (). Thus, in (figure 1(b)) the mechanisms are categorized as follow:
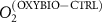
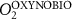
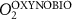
- (i)R0 indicates the dominance of AO dynamic transport over biological activity as a mechanism for the variation of O2,
- (ii)R1 indicates that transport and biogeochemical processes are of similar importance to the O2 changes. This suggests either a biological change concomitant with AO transport or transport of what result from remote biological response,
- (iii)R1 indicates the dominance of biological activity over dynamic transport to explain the variation of O2 consecutive to AO injection.

Along the coastlines with injection sites, in the Atlantic Ocean and across the North Pacific Equatorial Current, the correspondence is high (R0). In the Atlantic Ocean, the OMZ shrank even where it is devoid of AO stations (figure 1(b)). Advective transport is therefore the driving factor feeding AO into this OMZ. Since this OMZ is included in the North Atlantic Central Water (figure C2) and has its top located in the Subtropical Mode Water (figure 2(c)), it could benefit from an efficient supply of AO. This OMZ would therefore be fed by the small cell-dominated by meridional surface transport as well as by the upper limb of the Atlantic Meridional Overturning Circulation (AMOC, figure C2). An exact quantification of this would require a dedicated study. In the South Pacific Ocean and the Costa Rica Dome, a combined effect of biogeochemical feedbacks and dynamic transport (1) seems to explain the response to AO injection (figure 1(b)).
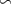
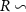
In the Indian Ocean, biogeochemical feedbacks dominate, especially in the BoB (1, figure 1(b)) where they appear to explain the counter-intuitive reduction of O2 concentration in the OXYBIO simulation with respect to CTRL (figure 2(b)) as treated in section 3.3. It is noteworthy that the different biogeochemical responses between the AS and BoB is likely due to different levels of oxygen deficiency [34].
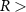
3.3. Impacts on nutrient recycling and biological activities
In the ocean, O2 is consumed by biological respiration, including OM remineralization. When the waters become suboxic, oxygen from NO3 is used for remineralization, a process called denitrification. OM remineralization is thus an internal source of 'recycled' fixed nitrogen (N) except in suboxic waters where denitrification is a sink of bioavailable nitrogen. In this section we investigate the potential cascading effects of the AO on the denitrification, remineralization, and Net Primary Production (NPP) (figures 3 and 4).

Figure 3. Changes after 100 years of AO injection (OXYBIO minus CTRL), of the annual mean of the (a) denitrification, (b) remineralization integrated on depth in %, (c) surface and (d) subsurface (between 200 and 600 m) nitrate concentration in . In (a) and (b) the orange line indicates the maximum extent of the area where denitrification occurs, and which corresponds to minimum denitrification. In (b) the red (blue) line is the increase (decrease) of 1% of the remineralization.
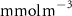
Download figure:
Standard image High-resolution image
Figure 4. The Net Primary Production integrated over the first 100 m is shown in (a)–(b). On the left, the NPP annual average of the last simulated year of the CTRL run, and in grey dotted line, is represented the Chlorophyll concentration of which defines the oligotrophic zones. On the right, the changes after 100 years of AO injection (OXYBIO minus CTRL) where the red (blue) line is the increase (decrease) of 1% of the NPP, and the black line demarcates the denitrification rise zone showed in figure 3(a).
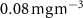
Download figure:
Standard image High-resolution imageDenitrification is absent in the Atlantic Ocean OMZ (figure C3) due to relatively greater natural ventilation than in the other OMZs. The decrease (up to 10% in the Pacific Ocean) or even total inhibition (in the BoB) of denitrification in response to AO injection (figure 3(a)) reduces the NO3 sink in initially (before AO injection) suboxic areas (figure 3, orange line). Note that at the surface, nutrients decrease except NO3 which increases (figure 3(c)), while at the subsurface all nutrients increase (figures C4, C5 and 3(d)). This suggests that the reduction of denitrification is likely to increase the availability of NO3 inducing the stimulation of NPP (figure 4(b)), thus increasing the exported production (figure 5(a)). As remineralization is constrained by OM [35–37], the additional supply of OM can thus amplify and maintain high remineralization leading to an increase in deep nutrient stocks [1]. Indeed, the seasonal thermocline of BoB is a major nutrient reservoir through the intense remineralization of exported OM [38]. However, by increasing NPP, nutrient uptake is amplified. Thus, the originally non-limiting nutrients may decrease until depletion [39] as in the BoB where NO3 limitation shifts to phosphate limitation for nanophytoplankton and changes from silicate to phosphate limitation for diatoms (figure C6).

Figure 5. Changes after 100 years of AO injection (OXYBIO minus CTRL). In (a) is the carbon export at 100 m (gC m−2 y−1), the green (magenta) line is the increase (decrease) of 1% of the carbon export. In (b) is the e-ratio (Export/NPP), the red (blue) line is the increase (decrease) of 1% of the ratio and in grey dotted line is represented the chlorophyll concentration of which defines the oligotrophic zones.
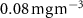
Download figure:
Standard image High-resolution imageIn conclusion, the expansion of OMZ indicates an imbalance between oxygen supply and biological O2 consumption, as in the case of BoB. Despite continuous oxygenated water injection and O2 transport through ocean dynamics, biological activity can still drive an expansion of an OMZ in some regions. The suppression of the nitrate sink (by inhibiting denitrification) and enhancing nutrient recycling (through remineralization) particularly at the seasonal thermocline is the cause. Moreover, the decline of the denitrification results in a decrease in nitrogen fixation (N2-fixation, roughly 50% in the vicinity of BoB) (figure C7). Landolfi's study [40] emphasized the vicious cycle induced by the spatial coupling of denitrification and N2-fixation. Due to stoichiometric imbalance, The NO3 loss through denitrification versus the nitrogen gain through the OM remineralization derived from N2-fixation leads to a net loss of fixed nitrogen that further stimulates N2-fixation. Counter-intuitively, the reduction in N2-fixation in the OXYBIO simulation, through the ihnibition of denitrification, could thus contribute to an increase in fixed nitrogen.
The other consequence particularly notable is the nonlocal environmental effects of AO injection. The AO stimulates biological activities beyond the zones of O2 change, especially in the Indian and tropical Pacific Oceans, except in the Northern Hemisphere where O2 changes and biological activities tally (figure 1(b)). While denitrification changes are mainly confined to the continental margins (figure 3(a)), remineralization changes are remotely triggered (figure 3(b)). Whether it is related to aerobic respiration (figure 3(b)) or NPP (figure 4(b)), biogeochemical changes follow the pathways of the advective currents (i.e. circulation of the subtropical gyres) (figures 1(b) and 4). In the Eastern Pacific Equatorial zone, phytoplankton is mainly Fe-limited in the CTRL simulation (figure C6). Thus, the gain in NO3 (figures 3(c) and (d)), due to reduced denitrification in this zone, would feed the initially NO3-limited phytoplankton communities in the areas surrounding the Pacific cold tongue and along the Central American coasts (figures C6(a) and (c)), through lateral surface transport. As a result, NPP increases north of the equator (figure 4(b)) along the Central American coasts (from the Gulf of Panama to the Mexican coast), where there are no injection stations (figure 1). This local NPP increase would trigger O2-consuming remineralization (figure 3(b)). Thus, the Costa Rica Dome, an eddy structure particularly influenced by the Costa Rica Coastal Current from the south, would be subject to exacerbated denitrification (up to 5%, figure 3(a)) due to this stimulation of remineralization bordering the dome. In the Tropical South Pacific Ocean, the edges of the denitrification zones (where it is lowest (figure C3), indicated by the orange line in figure 3(a)), show the largest decrease in denitrification, by 4%. Beyond these edges, the NPP remote response (rising to +3% off Chile (figure 4(b)) is triggered by the new supply of NO3 in zones that can be limited in NO3 (figure C6), which could explain the lack of NPP response near coasts where phytoplankton is mainly limited by Fe and Si (figure C6). Hence, these biogeochemical changes would be transported along the South Equatorial Current, feeding the subtropical gyre through the lateral transport [41].
Another striking region is the Indian Subtropical oligotrophic gyre (figure 4(a)), where the biological response is opposite to all other basins (figures 3(b) and 4). In BoB, the strong increase in NPP results in increased nutrient consumption (figure C4). This drop in nutrients could lead to a reduction in the supply of nutrients to the Indian Subtropical gyre through the South Java Current and explain the NPP decrease. In such a 'regenerative loop' regime [42] where most production is driven by recycled nutrients [43], the small reduction in OM remineralization may also explain the decrease in NPP in a chain-reaction.
3.4. Overview of the carbon cycle implications
Fertilization, whether natural [44] or artificial [45], can favor exported production through the development of ecosystems prone to exceptional blooms. These ecosystems are based on a biomass dominated by large cells (e.g. diatom [46–48]) and a NPP provided jointly by small and large cells [49]. The AO injection-induced denitrification inhibition does not appear to lead a change in the phytoplankton community despite NO3 enrichment (figure C8). Besides phytoplankton community composition, estimation of NPP is important for inferring the additional sequestered carbon. AO injection-induced NPP increase (+0.7%) is close to the magnitude of the decrease predicted by the low-emission scenario SSP1-2.6 (%) but is less than the decrease predicted by the high emission scenario SSP5-8.5 (%) by 2100 [11], which assumes a potentially negligible effect on the overall air–sea CO2 flux.
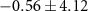
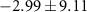
Because of the ability of artificial oxygenation to increase nitrogen concentration, it can also be suggested as an indirect means of fertilization by industry to stimulate the carbon pump. Yet, iron fertilization experiments revealed inconsistent export of production despite increased NPP [50]. In our study, despite the joint increase in NPP and exported production (figures 4(b) and 5(a)), the e-ratio decreases between 1%–14% depending on the region (figure 5(b)), which indicates a decrease in the efficiency of carbon export. Thus, due to a regionally variable duality between remineralization processes and OM export beyond injection sites, the effect of AO on carbon storage should not be overlooked in future studies.
3.5. Uncertainties regarding the AO injection impacts and the representation of biogeochemical processes
Several sources of uncertainty are possible when analyzing the effects of AO injection on the global ocean. The deployment scenario used in this study may represent one of them and may affect the results. For example, the choice of injection depth is constrained by structural robustness and cost. However, at 1000 m the oxygen utilization rate is 10 to 50 times lower than at 100 m [10, 51–54]. Hence, depth is an important causal parameter in the biogeochemical and ecosystem response to AO injection. The coarse resolution of the model used (2∘ resolution) prevents us from documenting technologies required to inject AO, which come under sub-grid processes but may affect AO efficiency [1, 15]. Yet, we can infer that AO should be undersaturated at the injection sites (figure C9). Thus, O2 saturation at injection sites (with no technical limitation taken into account) would allow the full dissolution of the AO. In addition, the spatial resolution implies process-related biases below the mesoscale spectrum [55]. Regional modeling of the AS highlighted the importance of eddy-induced advection on vertical and lateral O2 transport [56]. Increasing the resolution improves the explicit representation of eddy mixing that would be involved in the biased volume representation of OMZs in noneddy resolving ocean models. Thus, eddy mixing may be crucial in shaping marine habitats of oxygen-sensitive species [57].
Numerical modeling of OMZs and their ecosystems is an important socioeconomic issue because they are important fishing areas [58–60]. However, the current ocean microbiome modeling remains simple compared to the complexity that emerges from observations [61]. Also, AO injection showed that the evolution of carbon transfer to the deep ocean [62] as well as the fate of the OMZs, and consequently of the fisheries [58–60], require the analysis of phytoplankton and zooplankton communities. In our study, the microbial recycling mechanisms are parameterized lacking an explicit representation of bacterial communities despite the influence of O2 on controlling the availability of fixed nitrogen in the ocean [63, 64]. Regionally, explicit modeling of some ecosystems can improve the simulation of these processes, such as the diazotrophic community in the Tropical Pacific [65], especially since the anaerobic to aerobic transition in the ocean can cause strong negative feedback on nitrogen fixation [64]. Ultimately, ecosystem diversity differs according to the model used, and uncertainties in their applications remain [3, 61, 66, 67] and limit management or geoengineering decisions that are based solely on the use of models [68].
Despite these uncertainties, our results describe changes in biogeochemical processes in line with published regional studies on the effects of changing oxygen levels (e.g. in the Arabian Sea [57]). In the Pacific ocean, oxygenation through the modification of modeled vertical water exchange have also ultimately resulted in a deoxygenation and remote changes in biological activity [1]. However, given that biogeochemical processes tend towards a new equilibrium state after 20 to 40 years of simulation, as in BoB (not shown), we do not prejudge the effects of extending the experiment, nor of stopping the injection, either during or after the 100 year experiment, which could potentially lead to a global and regional new state.
4. Concluding remarks
Although model limitations must be kept in mind, our AO injection scenario reveals weak global oxygenation (+0.07%). Comparatively, CMIP models predict deoxygenation between 2.2% (SSP1-2.6 scenario), i.e. 30 times the oxygenation induced by AO, and 4.5% (SSP5-8.5 scenario) [11]. However, OMZs shrinkage (table 1) is in the lower limit of the OMZs expansion predicted by the RCP-8.5 scenario (between 1%–16% depending on the different CMIP5 models and thresholds [2]). Regionally, our study reveals deoxygenation and subsequent BoB OMZ expansion related to the dominance of biogeochemical processes, as well as non-local environmental effects in the Pacific and Indian Oceans, where biogeochemical and dynamic processes may be of similar importance. They are the result of the disruption of the nitrogen cycle through the inhibition of denitrification. The response of the Atlantic OMZ, devoid of denitrification, is marked by the AO transport allowing its shrinkage.
Given the uncertainties that may arise from both the scenario and the tools used, as discussed in section 3.5, regional and high-resolution studies are therefore needed to better understand the effects and risks of AO on benthic and pelagic ecosystems, higher trophic levels, and microbial communities, even on fisheries.
Acknowledgments
This work was supported by the Lhyfe SA company. We would like to thank Patricia Handmann and Stéphane Le Berre from Lhyfe for their contribution to this study. F Fransner was funded by the TRIATLAS European project (South and Tropical Atlantic climate-based marine ecosystem prediction for sustainable management; H2020 Grant Agreement No. 817578).
Data availability statement
The data that support the findings of this study are openly available at the following URL/DOI: https://doi.org/10.5281/zenodo.7377078. The data [69] are restricted by an access request.
Appendix A: Calculation of the correspondence
By analogy with the method used by Vinogradova et al 2007 and Bingham and Hughes 2008, we employed correspondence analysis to determine the extent to which anthropogenic O2 transport is involved in the variability of O2 over the 100 year injection period. Let be the evolution of the AO concentration during the 100 years of injection (O2 as a passive tracer), and the evolution of the O2 concentration during the 100 years of injection (i.e. oxygen varying with transport, biological activity, air-sea exchange, and in response to the AO injection). The correspondence is an adimentional value based on the estimation of the ratio between the variance of the difference between and and the variance of alone. Thus, we can compare whether:
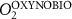

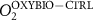
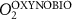
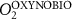
- (i)R0 indicates minor variation in biological activity ( - ) compared to dynamic transport (), as well as concomitant variation in and , revealing the preponderance of dynamic transport of AO over biological activity as a mechanism for the variation of O2;
- (ii)R1 indicates that the factors that differentiate from vary similarly to , so that biological activity varies similarly to the passive tracer transport. Thus, changes in are related to the transport of the biological activity outcome (such as organic matter (OM) and nutrients)
- (iii)R1 corresponds to a strong discrepancy between and , and the variation of biological activities is greater than that of dynamic transport. Consequently, the mechanisms that cause change are more related to biological activity than to dynamic transport, whether it is that of AO or that of biochemical elements. These various responses imply contrasted evolution of OMZs.

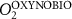
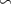
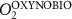
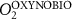
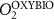
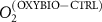
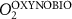
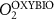
Appendix B: Definitions of water masses
The black solid lines in figures 2(c)–(f) show the isopycnes that delimit the water masses based on Talley et al 2011 and Pollard and Pu, 1985 (figure C2). In figure 2(c) Atlantic section at 2∘E, the potential density σ0 = 25 is the lower boundary to define the Subtropical Underwater (STUW), the North Atlantic Central Water is bounded between σ0 = 26.5 kg m−3 and 27.3 kg m−3 and including South Atlantic Central Water, between σ0 = 27.3–27.7 kg m−3 is the Intermediate Water. In (d) Pacific section at 120∘W, between σ0 = 24.5–26.5 kg m−3 it is the 'thermocline' which includes subtropical mode waters and between σ0 = 26.5–27.65 kg m−3 the 'intermediate waters' which includes subantarctic mode waters and North Pacific Intermediate Waters, and below is the 'deep ocean'. In (e) Bay of Bengal at 90 ∘E and (f) Arabian Sea at 70 ∘E, above σ0 = 26 kg m−3 it is the STUW, the Subtropical Mode Water is between σ0 = 26–26.8 kg m−3, below σ0 = 26.8 kg m−3 these are the 'intermediate waters' these are the intermediate waters which include, in the Arabian Sea, the Red Sea waters.
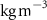
B.1. References of appendices A and B
Vinogradova NT, Ponte RM, Stammer D. 2007 Relation between sea level and bottom pressure and the vertical dependence of oceanic variability. Geophys Res Lett. 34. https://doi.org/10.1029/2006GL028588. Bingham RJ, Hughes CW. 2008 The relationship between sea-level and bottom pressure variability in an eddy permitting ocean model. Geophys Res Lett. 35. https://doi.org/10.1029/2007GL032662. Talley LD, Pickard GL, Emery WJ, Swift JH. 2011 Descriptive Physical Oceanography. Ed Elsevier. https://doi.org/10.1016/C2009-0-24322-4. Pollard, R. T., & Pu, S. 1985. Structure and circulation of the Upper Atlantic Ocean northeast of the Azores. Progress in Oceanography, 14(C), 443–462. https://doi.org/10.1016/0079-6611(85)90022-9.
Appendix C: Additional figures

Figure C1. The annual mean depth (m) of the upper limit of the Oxygen Minimum Zone of the threshold (a) above 600 m over the last simulated year of the CTRL run, (b) according to the climatology of WOA data, 2018 product [70].
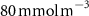
Download figure:
Standard image High-resolution image
Figure C2. Meridional section (zonal average) of the annual mean of the passive tracer (mmol m−3) after 100 years of AO injection (OXYNOBIO run) in the Atlantic Ocean. In dashed lines, the meridional overturning stramfunction in the classical latitude-depth coordinates. In blue (red), clockwise (counterclockwise) transport. In the Atlantic, the Atlantic Meridional Overturning Circulation (AMOC) cell is located above 3000 m and above the cell linked to the Antarctic bottom water (AABW). The Subtropical Underwater (STUW) framed in white, the North Atlantic Central Water (NACW), and South Atlantic Central Water (SACW) are described in the appendix B.
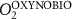
Download figure:
Standard image High-resolution image
Figure C3. Depth-integrated denitrification averaged over the last year of CTRL simulation. The red lines correspond to the isopleth of of O2 concentration between 200 and 600 m of depth (CTRL run).
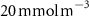
Download figure:
Standard image High-resolution image
Figure C4. Annual mean surface concentrations of phosphate (PO4), nitrate (NO3), silicate (SiO2), iron (Fe) over the last simulated year of the CTRL run (in and 'µmol m−3 for Fe) (a)–(d). Changes after 100 years of AO injection (OXYBIO-CTRL), of the annual mean surface concentrations of these nutrients (e)–(h).
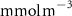
Download figure:
Standard image High-resolution image
Figure C5. Annual mean subsurface concentrations of phosphate (PO4), nitrate (NO3), silicate (SiO2), iron (Fe) over the last simulated year of the CTRL run (in and µmol m−3 for Fe) (a)–(d). Changes after 100 years of AO injection (OXYBIO-CTRL), of the annual mean subsurface concentrations of these nutrients (e)–(h).
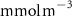
Download figure:
Standard image High-resolution image
Figure C6. Spatial distribution of each limiting nutrient (in brown the Phosphates, in purple the Iron, in cyan the Nitrates, and in red Silicate) of the surface nanophytoplankton (top) and diatoms (bottom) growth during the boreal (North Hemisphere) and Southern (South Hemisphere) summer of the last simulated year of the CTRL run (the left panels), and after 100 years of AO injection (right panels). Shades of opacity are given as an indicator of the level of total nutrient limitations. The limitation terms are defined according to [20].
Download figure:
Standard image High-resolution image
Figure C7. Annual mean N2 fixation rates (mmol N−2y−1) over the last simulated year of the CTRL run, and (b) changes after 100 years of AO injection (OXYBIO-CTRL).
Download figure:
Standard image High-resolution image
Figure C8. Relative abundance of diatoms in %, the left panel represents the last simulated year of the CTRL run, the right panel is the change after 100 years of AO injection (OXYBIO-CTRL) (the red solid line corresponds to +1%). In grey dotted line, is represented the Chlorophyll concentration of 0.08 mg m−3 which defines the oligotrophic zones.
Download figure:
Standard image High-resolution image
Figure C9. The left panel represents the annual mean of oxygen saturation at 150 m depth in CTRL simulation at the last simulated year. The right panel represents the percent AO saturation (the percent ratio of AO concentration to O2 saturation) at 150 m depth at each station after 100 years of AO injection at a rate of mol y−1.
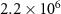
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Download figure:
Standard image High-resolution image{kind=link}