

Abstract
Drought-deciduous and evergreen species are both common in tropical forests, where there is the need to cope with water shortages during periodic dry spells and over the course of the dry season. Which phenological strategy is favored depends on the long-term balance of carbon costs and gains that leaf phenology imposes as a result of the alternation of wet and dry seasons and the unpredictability of rainfall events. This study integrates a stochastic eco-hydrological framework with key plant economy traits to derive the long-term average annual net carbon gain of trees exhibiting different phenological strategies in tropical forests. The average net carbon gain is used as a measure of fitness to assess which phenological strategies are more productive and more evolutionarily stable (i.e. not prone to invasion by species with a different strategy). The evergreen strategy results in a higher net carbon gain and more evolutionarily stable communities with increasing wet season lengths. Reductions in the length of the wet season or the total rainfall, as predicted under climate change scenarios, should promote a shift towards more drought-deciduous communities, with ensuing implications for ecosystem functioning.
Export citation and abstract BibTeX RIS
1. Introduction
Tropical vegetation must cope with water shortages during periodic dry spells in the wet season, and also over the course of the one-to-eight month long dry season that occurs in many tropical regions (Borchert 1999, Engelbrecht etal 2006, Feng etal 2013). During dry periods, soil water may be severely depleted by evapotranspiration, unless replenished through rainfall (if any). As soil water declines and air vapor pressure deficit increases, plants will regulate their internal water potential through reduction in transpiration rates; this physiological response is carried out through stomatal closure, which will simultaneously limit photosynthesis (Bucci etal 2005, Eamus and Prior 2001, Iida etal 2016, Wolfe etal 2016). Maintaining leaf area in spite of increasingly arid conditions imposes a carbon (C) cost on the plant, as maintenance respiration continues and the plant may sustain hydraulic damage. It might thus become advantageous from a C economy perspective for plants to shed leaves during the dry season (Givnish 2002, Manzoni etal 2015, Munne-Bosch and Alegre 2004). Drought-deciduous species would thus be expected to have a competitive advantage compared to evergreen species as the length of the dry season increases or total rainfall decreases (Givnish 2002, Manzoni etal 2015, Vico etal 2015).
Despite this first-order expectation, evergreen and deciduous species coexist in many tropical forests (e.g. Defries etal 2000, Enquist and Enquist 2011, Sobrado 1986, Williams etal 1997). This coexistence might arise due to the different ways in which physiological traits coordinate with phenological strategies, yielding comparable net C gain in the long term despite contrasting phenology. For example, evergreen species often exhibit deeper rooting depths than co-occurring drought-deciduous ones, allowing them to exploit a separate, more stable water source to support continued transpiration during dry periods (Hasselquist etal 2010, Iida etal 2016). In addition, deciduous species exhibit higher photosynthetic capacity, potentially supporting similar net C gain in the long-term relative to that of evergreen species with lower photosynthetic capacity; this is reinforced by the association of deciduous species with thinner leaves that levy a lower construction cost despite their shorter leaf life spans and more frequent turnover (Eamus and Prior 2001, Franco etal 2005). In the absence of such leverage, maintaining leaves during dry periods would require a combination of stable water sources, traits enabling tolerance to low leaf water potential, and C stores to sustain respiration with little net C assimilation. Leaf phenology and plant functional traits are also affected by plant nutrient availability, which may limit the number of leaves and photosynthetic machinery that can be supported (Aerts 1995, Bucci etal 2006, Ouédraogo etal 2016, Villagra etal 2013). Nutrient-limited conditions would thus favor evergreen species that typically exhibit lower leaf nitrogen (N) concentrations and lower nutrient requirements due to longer leaf longevity.
Moreover, in a plant community, each individual plant experiences the environmental conditions set by its surrounding community. If the C gain of an individual is markedly impacted by the surrounding soil water availability (as expected to be the case under water scarcity), then the fitness of such an individual is mediated by the water use habits of its neighbors. Thus, the water use patterns of a community affect the C balance of each individual, including that of a potential invasive individual with different traits (e.g. phenology; figure 1(a)), defining the evolutionary stability of the community (sensu Taylor and Jonker 1978). Hence, the relative advantage of specific functional traits and phenological strategies (intrinsic factors) for the C economy of individuals and communities is constrained by hydro-climatic drivers affecting soil water and by nutrient availability (extrinsic factors). The interactions of these intrinsic and extrinsic factors shape a complex plant fitness space, in which a range of trait combinations can be similarly successful in a given environment.

Figure 1 Schematic illustration of (a) plant phenology and (b) the hydrologic regimes. In (a), the different combinations of leaf phenological strategies considered are shown: EV and DD refer to homogeneous communities with evergreen or drought-deciduous strategies respectively; EV→DD (and DD→EV) refers to evergreen (and drought-deciduous) individuals invading a drought-deciduous (and evergreen) community. An example of soil moisture trajectory over a year is depicted in (b): soil moisture evolves randomly during the wet season and decays exponentially during the deterministic dry down in the dry season, starting from the random soil moisture s0 (the shaded curve is the probability density function of such random soil moisture). The exponential decay continues throughout the dry season in evergreen communities (dashed green line), while the soil moisture remains stable in drought-deciduous communities after leaves are shed and transpiration stops (solid red line).
Download figure:
Standard image High-resolution imageHere we evaluate the net C gain by plants adopting different phenological strategies (and associated traits), to quantify tree fitness in tropical forests and the evolutionary stability of tree communities, across gradients of extrinsic environmental drivers. By coupling eco-hydrological models with a minimalist plant C balance, average long-term net C gain (our proxy for fitness) is obtained as a function of rainfall statistics, N availability, and the plant functional traits of individual trees and the surrounding community. Tropical woodlands and savannas are described within a unified framework that accounts for the gamut of leaf phenological strategies and rainfall regimes in tropical seasonal and aseasonal climates. This framework is used to explore two complementary questions: (i) which leaf phenological strategy is the most productive in a given hydro-climatic regime? (ii) is that strategy evolutionarily stable or is invasion by individuals with a different leaf phenology possible?
2. Methods
To address these questions, a coupled model of plant community water and net C gain is developed. The following sections summarize how the soil water balance (section 2.1), plant C balance (section 2.2), and competition dynamics (section 2.3) are described. Model details and a list of symbols can be found in supplementary information (SI stacks.iop.org/ERL/12/065006/mmedia).
2.1. Soil water balance
We focus here on the soil layer hosting most of the roots (the rooting zone of Zr). At the stand scale, the dynamics of soil water over Zr are defined by the balance between input via rainfall and losses via evapotranspiration, surficial runoff and deep drainage (Porporato etal 2004, SI2). Rainfall occurs randomly at the daily scale. Two distinct hydro-climatic regimes are considered (figure 1(b)):
- A statistically stationary regime of duration Twet, with random rainfall events and dry spells. The frequency and duration of dry spells depend on the rainfall statistical properties and their interaction with plant water use. This wet regime corresponds to the wet season in seasonally dry climates; when it extends over the whole year, it describes climates without a pronounced seasonality in rainfall.
- An extended dry period, during which rainfall is assumed to be completely absent. This dry regime characterizes the dry season in seasonally dry climates. It has a prescribed duration of Tdry = 365-Twet days.
By considering climates with different predominance of these two regimes (i.e. different Tdry and Twet), the broad spectrum of climatic conditions of tropical forests and woodlands is explored, ranging from aseasonal climates (Twet ≅ 1 yr; note that these can be either wet or dry in nature depending on total rainfall), to markedly seasonal ones, with potentially long dry periods (large Tdry). Daily rainfall occurrence during the wet season is idealized as a Poisson process (with average frequency λ); the intensity of rainfall events is assumed to be exponentially distributed (with average depth α). Because the dry season is assumed to be devoid of rainfall, the total average annual rainfall is equal to the total average wet season rainfall, i.e. Rtot = αλTwet (Rodriguez-Iturbe and Porporato 2004).
2.2. Plant carbon balance
The fitness of a homogeneous monospecific stand is here quantified as the long-term average net C gain, 〈Gtot〉 (conceptually similar to Cowan 1986, Givnish 2002, Mäkelä etal 1996)—a realistic assumption in mature individuals where the relation between productivity and reproductive effort is generally monotonically increasing (Niklas and Enquist 2002). 〈Atot〉 is given by the difference between the net C assimilation during the wet and dry seasons (photosynthetic rate minus leaf maintenance and, in general, xylem repair C costs; 〈Atot〉) and the C costs of leaf construction, 〈Cc〉:
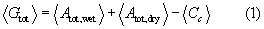
where 〈·〉 indicates ensemble averaging. All the terms on the right hand side are influenced by soil water availability and plant functional traits (SI2.2). Among these traits, we explicitly account for the maximum photosynthetic capacity (Amax) and leaf respiration rates (Rdark, Rlight); specific leaf area (SLA), leaf area index (LAI), and leaf longevity (LL); the maximum cost of xylem repair after embolism per unit leaf area (Rx); and plant water use thresholds (soil moisture levels below which photosynthesis is reduced and then completely ceased,  and sw respectively; for drought-deciduous species, also the soil moisture level corresponding to leaf shedding,
and sw respectively; for drought-deciduous species, also the soil moisture level corresponding to leaf shedding,  ). Most of these traits are linked by scaling relations (Wright etal 2004) and are associated with specific leaf phenological strategies and N available to leaves (SI2.3). In general, evergreen species have a lower photosynthetic capacity, SLA and leaf N concentration per unit leaf area than drought-deciduous species (figure S2; Eamus and Prichard 1998, van der Sande etal 2016, Vico etal 2015), which allows evergreens to achieve a higher LAI for a given N supply compared to drought-deciduous species (in agreement with observations; Asner etal 2003, Hély etal 2006, Myneni etal 1997).
). Most of these traits are linked by scaling relations (Wright etal 2004) and are associated with specific leaf phenological strategies and N available to leaves (SI2.3). In general, evergreen species have a lower photosynthetic capacity, SLA and leaf N concentration per unit leaf area than drought-deciduous species (figure S2; Eamus and Prichard 1998, van der Sande etal 2016, Vico etal 2015), which allows evergreens to achieve a higher LAI for a given N supply compared to drought-deciduous species (in agreement with observations; Asner etal 2003, Hély etal 2006, Myneni etal 1997).
The plant phenological strategy, functional traits and N and C economy dictate the temporal evolution of transpiration, photosynthesis and leaf area in response to soil water availability. In turn, the plant response to water shortage (reduction of transpiration and changes in leaf area) impacts the temporal evolution of soil moisture in the rooting zone by mediating the water fluxes out of the soil. Therefore, the phenological strategy indirectly controls the duration of plant water stress through its feedback on soil water availability. The actual leaf duration is not defined a priori, but rather it is the result of the interplay between soil water availability and use patterns, i.e. it is an emergent property of the climate-soil-vegetation system. In turn, the leaf phenology affects C assimilation directly, by constraining the length of the photosynthetically active periods, and indirectly because of the correlation between leaf phenology and other functional traits (e.g. Amax, Rdark, SLA).
The approach proposed here accounts in full for the randomness of rainfall occurrence during the wet season. Hence, soil moisture, photosynthesis and net C gain are to be treated as random variables. Their probability distributions and the long-term average of net C gain, 〈Gtot〉, are obtained analytically in SI3.
2.3. Species performance and invasibility
We first compare the fitness of homogeneous communities (either EV or DD in figure 1(a)) by considering which leaf phenology results in the highest productivity under a certain hydro-climatic forcing. Second, we identify evolutionarily stable leaf phenological strategies (combinations EV→DD and DD→EV in figure 1(a)), with water availability as the main driver of C balance. At the stand scale, the water use patterns of the existing community influences the C balance of the invading individual of different leaf phenology and thus may shape the likelihood of invasion (i.e. the evolutionary stability of a phenological strategy). A phenologically homogeneous community is considered stable when its fitness exceeds the fitness of the potential invader with a contrasting phenology, subjected to the environment set by the existing community (Parker and Smith 1990, Taylor and Jonker 1978). For consistency, the invader C balance is still expressed on a per unit area basis by equation (1).
In this framework, plant fitness and community evolutionary stability are interpreted over the long-term (e.g. on timescales commensurate with the lifespan of individual plants), as rainfall frequency, intensity, and the duration of the dry season are inherently unpredictable on shorter timescales.
3. Results
3.1. Phenological controls on community productivity
Figure 2 illustrates how the fitness of evergreen and drought-deciduous species is impacted by wet season duration (Twet) and total annual rainfall (Rtot). Regardless of phenological strategy, the net C gain increases and eventually saturates with increasing Rtot. In contrast, lengthening the wet season results in higher plant C gain for locations where the wet season durations are short (left side in each panel of figure 2), but does not alter C gain in evergreen species or it decreases it in drought-deciduous species towards aseasonal climates (right side).
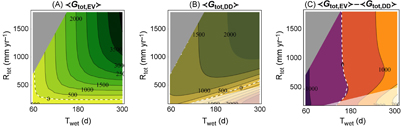
Figure 2 Effects of climatic conditions on annual average net C gain per unit ground area, 〈Gtot〉 (gC yr−1 m−2) (equation (1)) for (a) evergreen species and (b) drought-deciduous species, and (c) on the difference between the two strategies (evergreen species perform better when values are larger than 0), as a function of length of the wet season (Twet) and total annual rainfall (Rtot, varied by changing rainfall frequency at constant average rainfall event depth, α = 15 mm). All other parameters are as in table S2. The gray areas correspond to rainfall frequencies that are unrealistically high; the shaded areas refer to actual leaf durations in drought-deciduous species shorter than 75 d. In (a) and (b), climatic conditions to the right of and above the dashed white lines correspond to positive 〈Gtot〉, whereas in (c) conditions to the right of the dashed white line correspond to evergreen species performing better than drought-deciduous ones.
Download figure:
Standard image High-resolution imageThese contrasting effects stem from the underlying patterns in soil water availability. For given total annual rainfall Rtot and mean rainfall depth α, rainfall occurs more frequently over a shorter wet season, resulting in larger water losses via surface runoff and deep percolation, particularly at high Rtot. As Twet increases, rainfall events become less frequent, allowing at first lower runoff and more efficient water retention in soils and water use by plants. However, further lengthening of the wet season at fixed Rtot leads to a higher likelihood of dry spells, reducing water availability and C gain also during the wet season. Thus, at large Twet and low Rtot leaf shedding and flushing by drought-deciduous species becomes more frequent, limiting the effective leaf duration and causing soaring leaf construction costs (bottom right of figure 2(b)). Hence, in drought-deciduous species, a maximum in net C gain is observed at intermediate wet season durations for a given total annual rainfall; this optimal duration increases with Rtot. No optimal Twet emerges in evergreen species that are assumed to keep their leaves throughout the year. When comparing the two leaf phenological strategies (figure 2(c); conditions to the right of the dashed white line correspond to evergreen phenologies being more effective), leaf shedding emerges as the more productive strategy for Twet < 5 months, due to lower maintenance costs when photosynthesis is low.
As an example, we applied the model to two geographical transects: a west-east transect in South America (figure 3 left) and a south-north transect in Africa (figure 3 right). The estimated rainfall parameters summarize the local hydro-climatic regime (figures 3(a)−(d)): the decline in total annual rainfall along the South American transect is caused jointly by a decline in rainfall frequency and wet season length. Conversely, the length of the wet season increases markedly from south to north along the African transect and determines the rainfall trend there. When driven by these rainfall parameters, the model predicts higher average net C gain for evergreen species than drought-deciduous ones at the western and northern ends of the South American and African transects, respectively (figures 3(e) and (f)). The most productive leaf phenological strategy is expected to be dominant in the long term: indeed, model predictions of the most productive strategy are in good agreement with satellite observations (figures 3(g) and (h)).
Figure 3 Observed hydrologic regimes (a) and (b): average rainfall event depth, α, and average rainfall frequency, λ during the wet season; (c) and (d): total annual rainfall, Rtot, and duration of the wet season, Twet, (e) and (f) model estimates of evergreen and drought-deciduous net C gain and (g) and (h) observed fraction of evergreen vegetation (symbols) and comparison of the latter with model estimates of the most productive leaf phenological strategy (evergreen in dark green; drought-deciduous in light brown) along two transects: on the left, a west-east transect in South America (at latitude −7.25°); on the right, a south-north transect in Africa (at longitude 22.75°). Rainfall parameters were obtained from TRMM data (Huffman et al 2010) for the period 1998–2015; vegetation cover from Defries et al (2000) (see details in SI4). Model estimates are based on the same plant traits as in figures 2(d)−(f), but with rainfall parameters (α, λ and Twet) from the TRMM data. Note the difference in the scale of the ordinates between (e) and (f).
Download figure:
Standard image High-resolution imageAlso plant functional traits affect the annual average net C gain, 〈Gtot〉. We focus here on two traits that are known to vary markedly among evergreen species (rooting depth, Zr,EV, and leaf longevity, LLEV), while keeping the traits of the drought-deciduous species constant. Figure 4 summarizes the isoclines corresponding to the same 〈Gtot〉 for drought-deciduous and evergreen communities for different rainfall regimes and plant traits. Trait combinations for the evergreen species above each curve lead to homogenous evergreen community having a higher fitness that the deciduous one, under the specified rainfall pattern. Longer wet seasons (and less frequent rainfall events; lighter color shades) are beneficial for evergreen species (curves shifting downwards). In addition, total rainfall amount does not markedly influence the most beneficial phenological strategy, except under longer wet seasons or very shallow rooting depths. Longer leaf duration counters the disadvantage of having shallower roots in evergreen species, because it reduces leaf construction costs and allows for a more efficient use of nutrients, despite lower photosynthetic capacity (SI2.3). Deeper roots for the evergreen species reduce the need to have long lived leaves to achieve a net advantage over drought-deciduous species. This is suggested by the inverse relationship between LLEV and Zr,EV along the contour lines. The compensating effect of long lived leaves for shallower roots becomes stronger when considering climates characterized by shorter wet seasons (darker lines), because only increased access to water storages can sustain comparable plant activity during the prolonged dry seasons. Under those conditions, only evergreen species with deep roots (and durable leaves) can be more productive than drought-deciduous ones.

Figure 4 Evergreen trait combinations (rooting depth, Zr,EV, and leaf longevity, LLEV) for which evergreen and drought-deciduous species have the same average annual C gain 〈Gtot〉, for different rainfall totals (solid lines, Rtot = 1200 mm; dashed lines, Rtot = 600 mm) and different rainfall frequencies and durations of the wet season (shades; climatic parameters are reported in the legend; in all cases, α = 12 mm). The plot region above each curve contains the trait combinations of the evergreen species for which a homogeneous evergreen community has a higher productivity than the drought-deciduous one. All the other parameters are as in table S2.
Download figure:
Standard image High-resolution image3.2. Evolutionary stability of plant communities
For set plant traits, the climate space can be divided into three regions on the basis of which phenological strategy is most productive in a homogeneous community and whether such community is evolutionarily stable (figure 5(a)). The length of the wet season plays a crucial role. Short wet seasons result in higher productivity of drought-deciduous communities (regions a and b). Such communities could be invaded by evergreen individuals when wet seasons are of intermediate length (region b), because evergreen species can exploit the soil water remaining after drought-deciduous species shed their leaves. Even longer wet seasons (region c) result in evergreen communities being not only more productive than drought-deciduous ones but also evolutionarily stable. Rainfall amount (and hence frequency) plays a role in defining the most productive and evolutionarily stable community only when Rtot ≲ 600 mm. There, a further decrease in rainfall first lengthens then shortens the wet seasons for which a drought-deciduous community is evolutionarily stable (region a) or more productive than an evergreen community (region b). An initial increase in Twet for a given Rtot decreases runoff by distributing rainfall more uniformly, allowing a more effective water use during the wet season. However, longer wet seasons when Rtot is low are characterized by rare rainfall events and more frequent dry spells also during the wet season, leading to leaf shedding in the drought-deciduous species, with the associated C cost of the subsequent leaf flushing.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 5 Partition of the (a) climate and (b) plant trait parameter space into different regions, based on productivity and evolutionarily stability criteria. The climatic parameters considered are total annual rainfall, Rtot, and length of the wet season, Twet; the plant traits are the leaf longevity for evergreen species, LLEV, and soil moisture threshold for leaf shedding, , for drought-deciduous species. The parameter space comprises three distinct regions: (a) stable drought-deciduous communities (i.e. an evergreen individual cannot invade), which also exhibit a productivity higher than a homogeneous evergreen one; (b) conditions under which a drought-deciduous homogeneous community is more productive than but can be invaded by an evergreen one; (c) stable and more productive evergreen than drought-deciduous community. Parameter values are summarized in table S2. In (a), the gray area corresponds to unrealistically high rainfall frequencies; the shaded area refers to actual leaf durations in drought-deciduous species shorter than 75 d.
Download figure:
Standard image High-resolution image{kind=link}
Also plant functional traits affect the productivity and evolutionary stability of drought-deciduous and evergreen communities, for a given rainfall regime. Both parameters considered here have potentially contrasting effects on C economy. Longer leaf duration for evergreen species reduces the costs of leaf construction and the respiration rate, but it is also associated with lower maximum photosynthetic capacity per unit leaf area and larger leaf area (SI2.3). Drought-deciduous species shed their leaves when soil moisture reaches the threshold : a higher threshold potentially reduces the net C gain and enhances the frequency of leaf exchange. The net effects of these changes are represented in figure 5(b). Evergreen species are more productive and evolutionarily stable for large LLEV or , while drought-deciduous species are advantaged in the opposite case. At intermediate parameter values, a drought-deciduous community is more productive than an evergreen one, but not evolutionarily stable.
While the results above do not account for any C cost of embolism repair, our analyses suggest that the general patterns regarding fitness (figures 2 and 4) and evolutionary stability (figure 5) hold if plants are also faced with the costs of embolism repair due to drought-induced hydraulic impairment (Anderegg etal 2016, Barigah etal 2013). The larger this cost is, the more pronounced the shift to the right of the boundaries among the different regions (see figures S7–S9), indicating that drought-deciduous species will become increasingly favored thanks to the cavitation prevention resulting from leaf shedding.
4. Discussion and conclusions
4.1. Theoretical advances and limitations
The approach developed here unifies eco-hydrological and C economy models with plant trait relations for tropical forests, providing a measure of not only plant fitness but also evolutionary stability of communities, in aseasonal to markedly seasonal climates. The approach builds on earlier studies exploiting a cost-benefit approach to leaf C economy (Chabot and Hicks 1982, Givnish 2002, Manzoni etal 2015, Vico etal 2015), which recently received renewed support (Buitenwerf and Higgins 2016). The main novelty rests on (i) the explicit and detailed accounting for total annual rainfall, its distribution during the year, and their inherent unpredictability, which play a crucial role in tropical ecosystems (Guan etal 2014); and (ii) a simple, yet mechanistic description of the traits and N economy associated to leaf phenology—an aspect previously invoked to explain observed phenological patterns (Chabot and Hicks 1982, Givnish 2002) but seldom included in mechanistic models (see Weng etal 2017 for an example relative to boreal temperate forests). Differently from previous numerical approaches (Hély etal 2006, Vico etal 2015), closed analytical formulas for the fitness proxy, 〈Gtot〉, are obtained, thus facilitating the exploration of the effects of hydro-climatic drivers and plant traits. The advances presented here extend recent stochastic eco-hydrological models (Dralle and Thompson 2016, Feng etal 2012, Viola etal 2008), by incorporating leaf flush and shed dynamics. Our results well describe observed predominance of specific leaf phenologies across two transects (figure 3), by accounting for both extrinsic (hydroclimatic) and intrinsic factors (plant traits and phenological strategies), despite being less detailed than recent contributions that include plant hydraulic controls on stomatal conductance, process-based photosynthesis sub-models, and more complex, trait-based parameterizations (Fatichi etal 2016, Manzoni etal 2015, Xu etal 2016).
Compared to most existing eco-hydrological models (with few recent exceptions; see Farrior etal 2015, Weng etal 2017), the inclusion of an evolutionary stability analysis in the seasonally dry context adds a further level of interpretation of results and provides an explanation for coexisting leaf phenological strategies. Specifically, the evolutionary stability analysis provides the conditions under which species with different leaf phenological strategies can exploit niches in otherwise uniform communities. This approach combines the concepts of fitness and niche differences between community and invader (MacDougall etal 2009) and identifies the potential for mixed communities, but not necessarily the success of invasive alien species (sensu Richardson and Pyšek 2006), in particular in combination with other disturbances.
The simplifications adopted here caution against the indiscriminate application of this model. From a hydrologic perspective, the choice of pursuing an analytical approach lead to neglecting the transient rewetting at the beginning of the wet season and any rainfall during the dry season. From a plant eco-physiological perspective, the proposed model does not include a full trait-based description of plant activity and does not account for additional limiting factors (e.g. light availability). Nevertheless, it has the advantage of a concise representation that is useful for assessing broad-scale eco-hydrological patterns (as in Kumagai and Porporato 2012), while still providing the mechanistic understanding necessary to predict future responses of tropical forests (Corlett 2016). Furthermore, information about the effects of some of the mechanisms neglected here could still be inferred from the proposed approach. For example, plant water storage—typical of succulent plants—would in principle have the same effects as a deeper rooting zone, i.e. it would mostly be beneficial for evergreen species, particularly in drier climates (not shown).
Leaf flushing and shedding are here controlled by a pre-defined moisture threshold. Other existing schemes are based on hypothesized controls via photosynthesis or leaf water potential and turgor pressure (Arora and Boer 2005, Manzoni etal 2015, Xu etal 2016), but all share the same outcome—leaves are flushed when moisture increases and shed when moisture decreases. In reality, leaf phenology cannot perfectly track moisture conditions fluctuating around the threshold for leaf abscission, so that the predicted construction costs should be considered as an upper limit, particularly under low rainfall totals spread over a long wet season (shaded areas in figures 2 and 5). Furthermore, while phenology is typically linked to soil moisture availability in tropical dry forests (Borchert etal 2002, Lima and Rodal 2010, Wolfe etal 2016), some species follow other environmental clues (e.g. photoperiod) and flush leaves during dry periods, possibly tapping deep water stores (Borchert 1994). By focusing on species sensitive to soil moisture dynamics only ('opportunistic' species), our description may underestimate the net C gain in those drought-deciduous species that flush their leaves before the end of the dry season in anticipation of the wet season (i.e. 'scheduled' leaf flushers; Borchert etal 2002, Vico etal 2015). The next challenge for seasonally dry ecosystem models is to incorporate these moisture-independent phenological controls into a coherent modeling framework.
Finally, the evolutionary stability provides an objective criterion for the likelihood that a species could effectively exploit excess water resources in the existing community with different phenology, thus invading it. A lack of evolutionary stability does not necessarily translate to the community being successfully supplanted by the invading species (MacDougall etal 2009), rather here we interpret evolutionarily unstable communities as those with enhanced potential of hosting a more phenologically diverse community. In fact, in the community stability analysis, we assumed that the invaders experience the environment of the existing community (as in Parker and Smith 1990, Taylor and Jonker 1978)—a condition valid when the abundance of the invasive species is low. Furthermore, when fitness is determined in large part by water limitations, this assumption is reasonable in the case of drought-deciduous species entering an evergreen community, because the leaf shedding of the invader provides a negligible advantage in water availability to the whole community. In the opposite case of an invasive evergreen, our model stipulates that soil water remains constant after deciduous species shed their leaves (bare soil water evaporation is neglected). Because water losses from the soil and the evergreen plant are expected even after the leaves of the community are shed, water availability for the invading evergreen may be overestimated towards the end of the dry season. The predicted region of invasion by evergreen species may thus be larger than it would be had we included residual water losses in the deciduous community. Moreover, a drought-deciduous individual invading an evergreen community may experience limiting factors not related to water availability, such as light limitations, thus making the proposed approach less suitable for e.g. the case of a dense evergreen forest. We note however that our results point to evergreen communities being evolutionarily stable also with reference to water availability; light limitation would further strengthen their stability, thus not altering the broad patterns of our predictions.
4.2. Consequences of climate change for tropical forests
In agreement with more complex numerical models (Guan etal 2014, Hély etal 2006), our results suggests that the length of the wet season is the most relevant climatic determinant of net C gain, the most beneficial phenological strategy, and its evolutionary stability (figure 2). For drought-deciduous species under set total rainfall, an intermediate length of the wet season emerges as the one associated with the maximum net C gain, as already shown by a simpler model (Feng etal 2012) and remote sensing (Souza etal 2016). Rainfall totals and rainfall pattern within the wet season (as summarized by the rainfall frequency and event depth) play a less prominent role, with the exclusion of extremely arid regions (figures 2–4). To our knowledge, no empirical results are available in support of these results, as rainfall manipulation experiments have been performed only in wet tropical forests and in temperate regions (Allen etal 2017).
Predicted changes in rainfall amount and seasonality over the tropics are still uncertain (Chadwick etal 2016), although tropical regions are expected to experience a decrease in the overall rainfall amount along with a lengthening of the dry season (Fu etal 2013, Pascale etal 2016). According to our model predictions, the lengthening of the dry season alone (a shift to the left in figure 5) would in general be beneficial for drought-deciduous species, potentially transitioning evergreen stable communities to more productive (but not stable) drought-deciduous communities, and unstable drought-deciduous communities to stable ones, in agreement with the results of more complex models (Alo and Wang 2008). Under current climates, moving along aridity gradients indeed leads to an increased prevalence of drought-deciduous species as the wet season becomes shorter and rainfall decreases (Enquist and Enquist 2011, Fauset etal 2012, Feeley etal 2011, Ouédraogo etal 2016). This pattern is not expected to hold in currently dry regions, where a reduction in rainfall amount may even have the opposite effect (figure 5(a)), if wet season length would remain the same, or no effect at all, should the wet season become shorter.
Our results also suggest that changes in species composition will be mediated by the traits associated with each phenological strategy. Drought-deciduous species shedding leaves at lower soil moisture values can be more productive than those shedding leaves at higher moisture thresholds (figure 5(b)). Lower soil moisture threshold for leaf shedding may lead to shorter leafless periods at the end of the dry season. This 'brevi-deciduous' strategy can thus become more common where water availability decreases. In contrast, evergreen species greatly benefit from leaves with longer durations, in particular when roots are shallow (figure 4), despite the lower C assimilation potential of more durable leaves.
In conclusion, we developed a stochastic framework that accounts for the role of daily and seasonal rainfall patterns on the C economy of deciduous or evergreen species over the gamut of tropical climates, with plant traits linked to leaf phenological strategy.We have underscored the importance of considering not only the overall rainfall amount but also its intra-annual variability in defining the productivity and stability of phenological strategies in a given hydro-climatic regime. Understanding the potential shifts in species composition of tropical forests in response to future changes in these daily and seasonal rainfall patterns will allow us to anticipate the ensuing feedback to ecosystem functioning.
Acknowledgments
GV gratefully acknowledges the support of the project 'TC4F—Trees and Crops for the Future' funded through the Swedish government's Strategic Research Environment 'Sustainable use of Natural Resources' and of the Swedish Research Council (Vetenskapsrådet) under grant 2016–04910; SM was supported by the Bolin Centre for Climate Research, Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS) under grant 2016–00998, and the US National Science Foundation under grant FESD-1338694; XF thanks the funding from the NOAA Climate and Global Change Postdoctoral Fellowship; DD and SET acknowledge funds from the National Science Foundation under grant EAR-13311940 for the Eel River Critical Zone Observatory; SET was also partially supported by the National Science Foundation under grants IOS-1441396 and IOS-1457400.