

Abstract
Vegetation composition shifts, and in particular, shrub expansion across the Arctic tundra are some of the most important and widely observed responses of high-latitude ecosystems to rapid climate warming. These changes in vegetation potentially alter ecosystem carbon balances by affecting a complex set of soil–plant–atmosphere interactions. In this review, we synthesize the literature on (a) observed shrub expansion, (b) key climatic and environmental controls and mechanisms that affect shrub expansion, (c) impacts of shrub expansion on ecosystem carbon balance, and (d) research gaps and future directions to improve process representations in land models. A broad range of evidence, including in-situ observations, warming experiments, and remotely sensed vegetation indices have shown increases in growth and abundance of woody plants, particularly tall deciduous shrubs, and advancing shrublines across the circumpolar Arctic. This recent shrub expansion is affected by several interacting factors including climate warming, accelerated nutrient cycling, changing disturbance regimes, and local variation in topography and hydrology. Under warmer conditions, tall deciduous shrubs can be more competitive than other plant functional types in tundra ecosystems because of their taller maximum canopy heights and often dense canopy structure. Competitive abilities of tall deciduous shrubs vs herbaceous plants are also controlled by variation in traits that affect carbon and nutrient investments and retention strategies in leaves, stems, and roots. Overall, shrub expansion may affect tundra carbon balances by enhancing ecosystem carbon uptake and altering ecosystem respiration, and through complex feedback mechanisms that affect snowpack dynamics, permafrost degradation, surface energy balance, and litter inputs. Observed and projected tall deciduous shrub expansion and the subsequent effects on surface energy and carbon balances may alter feedbacks to the climate system. Land models, including those integrated in Earth System Models, need to account for differences in plant traits that control competitive interactions to accurately predict decadal- to centennial-scale tundra vegetation and carbon dynamics.
Export citation and abstract BibTeX RIS
1. Introduction
Northern high-latitude regions have experienced rapid warming in recent decades (Berner and Heal 2005, IPCC 2013). Ecosystems are responding to this warming in ways that may exacerbate or slow climate change, through changes in vegetation and processes influencing the thaw of permafrost, which stores about twice as much carbon as does the current atmosphere (Ping et al 2008, Schuur et al 2008, Hugelius et al 2014). Several lines of observational evidence indicate that rapid climate warming over the past few decades has resulted in (a) shifts in phenology (Myneni et al 1997, Tucker et al 2001, Verbyla 2008, McManus et al 2012, Prevéy et al 2019); (b) thawing of permafrost (Brown and Romanovsky 2008, Schuur et al 2015, Hugelius et al 2020); (c) thermokarst development (Schuur et al 2007, Jones et al 2015, Turetsky et al 2020); (d) more frequent and intense wildfire events (Flannigan et al 2009, IPCC 2013); and (e) alteration of landscape thermal dynamics, hydrological (Liljedahl et al 2016, Teufel and Sushama 2019), and nutrient cycling (Xue et al 2016, Sarneel et al 2020). These changes all contribute to a complex set of soil–plant–atmosphere interactions, potentially altering ecosystem carbon balances (Weintraub and Schimel 2005, Hudson et al 2011, Sistla et al 2013, Ravn et al 2020).
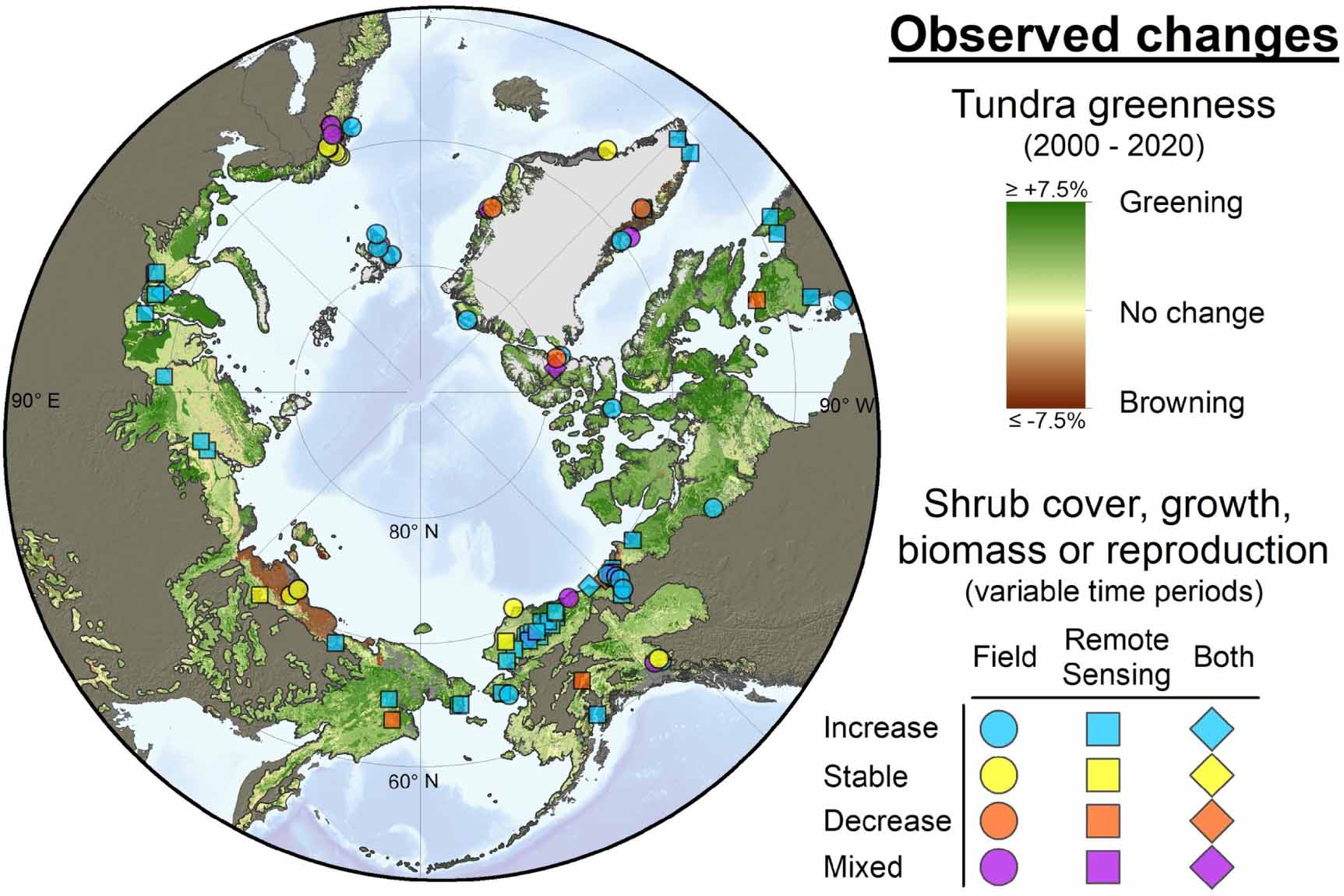
Rapid climate warming in northern ecosystems may also drive changes in competitive interactions and thereby alter plant species composition and abundance (Shaver et al 2000, Pieper et al 2011, Cahoon et al 2012, Elmendorf et al 2012a). In this review, we focus on the Arctic tundra, which is an ecosystem with diverse plant functional types (PFTs, plant groups with similar function and forms, e.g. evergreen shrubs, deciduous shrubs, graminoids, forbs, non-vascular vegetation) co-existing across several bioclimatic subzones (Walker et al 2005). Historical vegetation distributions reconstructed from paleo-records indicate a higher relative abundance of shrubs in the High Arctic and a more northern treeline during a warmer mid-Holocene compared to the present (Bigelow et al 2003). Across much of this region, recent changes in vegetation composition (e.g. observed increases in woody plant (mainly tall deciduous shrubs) growth, distribution, and relative abundance; also known as Arctic shrubification) have been reported (Myers-Smith et al 2011a, Elmendorf et al 2012b, García Criado et al 2020). We note that most observations of recent tundra shrub expansion mainly refer to the expansion of tall deciduous shrubs. Shrub abundance and changes inferred from remote sensing were shown to exhibit spatial heterogeneity, with greater observed increases in the Low Arctic (Lantz et al 2010, Berner et al 2020; figure 2).
Shrub growth has been shown to be climate sensitive (Forbes et al 2010, Myers-Smith et al 2015), and shrub increases are thought to be in response to climate warming (Cornelissen et al 2001, Wahren et al 2005, Walker et al 2006, Elmendorf et al 2012b, García Criado et al 2020), with other drivers (e.g. soil moisture, snow dynamics, disturbance, and herbivory) also playing roles (Martin et al 2017, Niittynen et al 2020). Increases in shrub growth have often occurred at the expense of non-vascular vegetation (e.g. lichens and bryophytes) (Cornelissen et al 2001, Elmendorf et al 2012a, Hollister et al 2015). These responses have been corroborated by a broad range of evidence, including in-situ observations (Hudson and Henry 2009, Callaghan et al 2011, Elmendorf et al 2012b), warming experiments (Walker et al 2006, Elmendorf et al 2012a), dendroecology (Forbes et al 2010, Myers-Smith et al 2015), repeat photography (Tape et al 2006), and satellite remote sensing (Forbes et al 2010, McManus et al 2012). This evidence strongly suggests that with recent climate warming shrubs have become more competitive across the Arctic tundra.
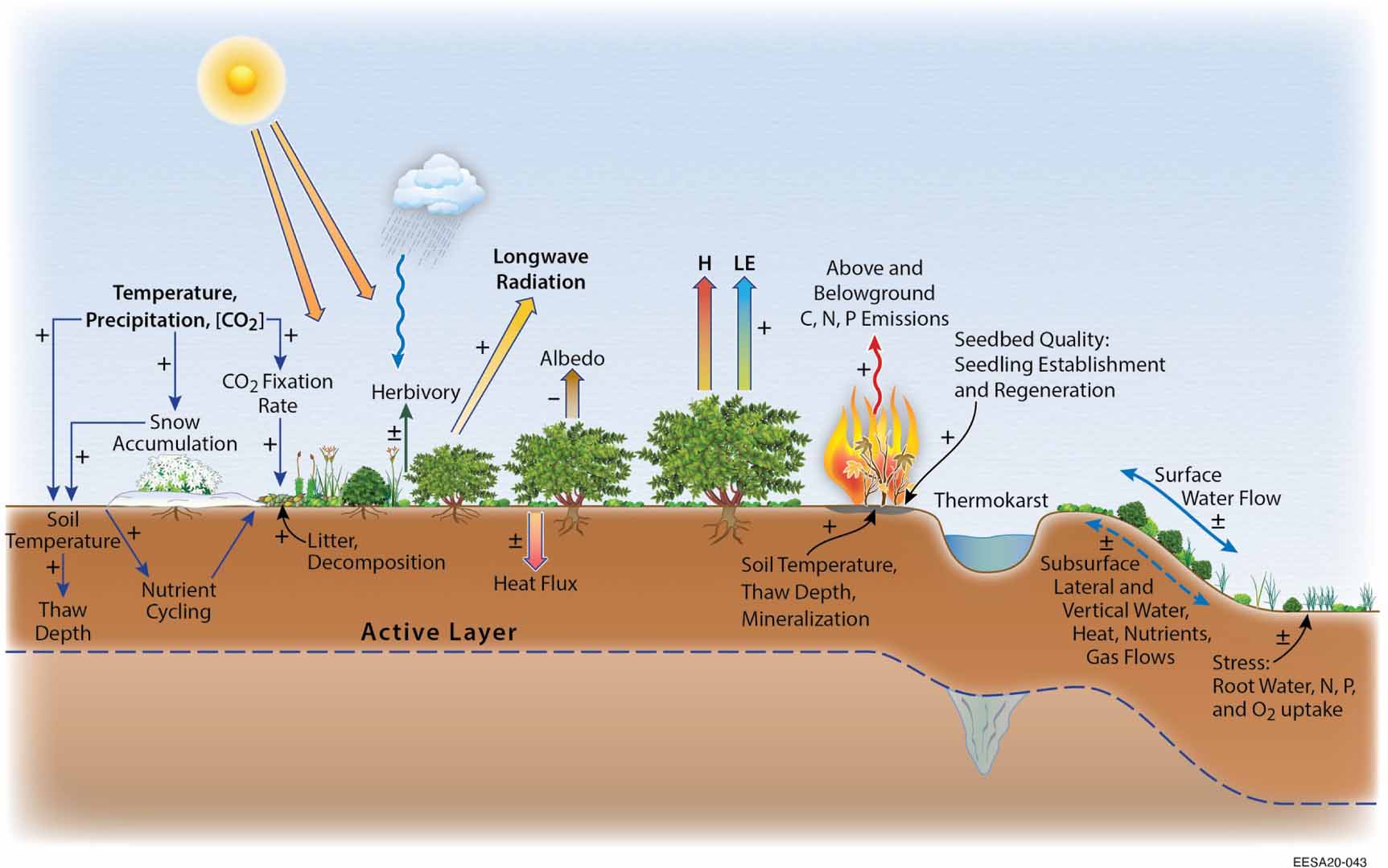
Under changing environmental and climatic conditions, several interacting processes affect Arctic tundra vegetation composition and carbon and nutrient cycling (figure 1). Competitive abilities of tundra plants are strongly controlled by differences in traits that control carbon and nutrient investments and retention strategies in leaves, stems, and roots; for example leaf nutrients and optical traits, root traits, plant hydraulics, morphological and phenological traits (Bjorkman et al 2018, Myers-Smith et al 2019a). Growth and expansion of shrubs affect the tundra carbon balance by enhancing ecosystem carbon uptake (Walker et al 2006, Forbes et al 2010, Elmendorf et al 2012a, Tremblay et al 2012) and by altering ecosystem respiration, which affects soil carbon stocks (Sistla et al 2013, DeMarco et al 2014a, Lynch et al 2018, Gagnon et al 2019, Ravn et al 2020) and nutrient cycling (DeMarco et al 2014b, Christiansen et al 2018a, Wang et al 2018, Prager et al 2020). Changes in net ecosystem carbon exchange driven by shrub expansion are also affected by complex feedback mechanisms, such as alteration of surface energy budgets (Chapin et al 2000, Blok et al 2010, Bonfils et al 2012, Lafleur and Humphreys 2018), snowpack dynamics (Liston et al 2002, Sturm et al 2005a, Marsh et al 2010, Myers-Smith and Hik 2013), and permafrost degradation (Blok et al 2010, Lawrence and Swenson 2011, Nauta et al 2015). Many of these shrub-driven changes, for example in energy balance and snowpack, may also affect climate; but in this review, we focus on the carbon-mediated feedback processes.

Figure 1. Schematic diagram that shows key climatic and environmental controls on tundra shrub expansion. The figure shows a complex set of soil–plant–atmosphere interactions driven by (1) climate change (increase in surface air temperature, precipitation and atmospheric (CO2)), (2) soil moisture and snow dynamics, (3) topography, (4) permafrost thaw, (5) nutrient dynamics, (6) disturbance (e.g. wildfire), and (7) herbivory, and interactions among these factors. These drivers may alter vegetation composition and lead to the expansion of shrubs. Shrub expansion and its effects on snowpack dynamics, litter inputs, permafrost degradation, and surface energy budgets may also affect net ecosystem carbon exchanges. Signs (±) on the arrows represent increases or decreases.
Download figure:
Standard image High-resolution imageAlteration of surface energy and carbon balance driven by changes in the abundance of woody shrubs may have important ecological and climatic implications. For instance, these implications include alteration of ecosystem structure, function, and feedbacks to climate (Loranty and Goetz 2012), fire fuel (Camac et al 2017), animal habitat (Tape et al 2016), and traditional gathering activities (Henry et al 2012). However, the mechanisms through which changes in climatic and environmental controls alter the composition of Arctic ecological communities remain unclear and the representations of these mechanisms in models remain incomplete.
Important goals in this review are to explore information from the literature using the Preferred Reporting Items for Systematic Reviews and Meta-Analyses approach (supplementary methods (available online at stacks.iop.org/ERL/16/053001/mmedia)) and to facilitate improved representations of tundra shrub processes in models used to assess carbon-climate feedbacks (Jones et al 2016). For example, land models are just beginning to include wildfire and its impacts on shrubs; the ability for shrub ranges to shift; and interactions among decomposition, nutrients, and shrub growth. Further, these land models do not include the effects of topographical changes due to permafrost thaw and thermokarst. As a result, models predict a wide range of carbon cycle responses over the 21st century, including positive (Qian et al 2010) and negative (Zhuang et al 2006) effects of future climate on ecosystem carbon stocks. Thus, predictions of the magnitude and direction of carbon-climate feedbacks associated with tundra shrub expansion remain uncertain in land models (Bonfils et al 2012, Druel et al 2019, Mekonnen et al 2018a, 2018b).
Decadal- to centennial-scale prediction of tundra carbon cycle dynamics requires land models that consider the wide array of ecological processes and their interactions and are robustly evaluated against observations. The paper is therefore organized into sections discussing (a) observed shrub expansion, (b) key climatic and environmental controls and mechanisms that affect the vegetation composition of Arctic tundra ecosystem, (c) overall impacts of shrub expansion and interacting feedback mechanisms that affect ecosystems carbon exchanges, and (d) observational research gaps and future directions for land models. Below we explore the literature and highlight the processes that are most relevant to each section.
2. Observed shrub expansion across the Arctic tundra
2.1. Satellite observations of Arctic greening
Multiple Earth-observing satellites provide evidence that summer normalized difference vegetation index (NDVI) widely increased (spectral greening) and locally decreased (spectral browning) during recent decades across the Arctic. Circum-Arctic spectral greening and browning trends have primarily been assessed during recent decades using coarse-resolution (∼8 km) summer NDVI derived from the Advanced Very High Resolution Radiometer (AVHRR) sensor (Myneni et al 1997, Bhatt et al 2010, Beck and Goetz 2011, Guay et al 2014, Andersen and Andreassen 2020, Myers-Smith et al 2020). However, higher-resolution NDVI data sets from the (500 m) MODIS (Guay et al 2014, Jenkins et al 2020, Myers-Smith et al 2020) and (30 m) Landsat (Berner et al 2020) satellites have been increasingly used for circum-Arctic assessments. A recent AVHRR NDVI analysis found spectral greening and browning across 38% and 3%, respectively, of the Arctic from 1982 to 2014, with greening evident over large parts of the eastern Eurasian and North American Low Arctic (Park et al 2016). Xu et al (2019) used AVHRR NDVI to assess controls on spring greenup and compared these controls with those inferred from MODIS (Xu et al 2018). Here we update the Landsat NDVI trend analysis from Berner et al (2020) to extend from 2000 to 2020 (instead of to 2016; supplementary material). We find spectral greening and browning across 27% and 8% of the Arctic from 2000 to 2020, respectively, along with a 3.9% increase in mean Arctic NDVI during this period (Mann–Kendall trend test: p = 6.6 × 10−6, τ = 0.65, n = 21 years; figure 2). While the magnitude and spatial patterns of spectral greening and browning differ somewhat among satellite NDVI data sets (Guay et al 2014), these products nevertheless show overall spectral greening of the Arctic during recent decades (Berner et al 2020, Jenkins et al 2020, Myers-Smith et al 2020). Arctic spectral greening has been linked with increasing summer air and soil temperatures, permafrost thaw, and loss of sea ice (Bhatt et al 2010, Keenan and Riley 2018, Berner et al 2020, Peng et al 2020), and with increasing cover and growth of shrubs and other vascular plants (e.g. Forbes et al 2010, Fraser et al 2011, Frost et al 2014, Andreu-Hayles et al 2020).

Figure 2. Observed changes in tundra greenness and shrub dynamics across the Arctic tundra biome. Changes in tundra greenness are based on trends in maximum summer NDVI (NDVImax) from 2000 to 2020 derived from Landsat satellite observations. Specifically, the map shows the percent change in mean NDVImax among sampling sites (n = 41 341) stratified by ecological land units (n = 186) with similar vegetation, soil, and climate. The trend map was modified and updated from Berner et al (2020). Observed changes in shrub cover, growth, biomass, and reproduction were based on a systematic literature review updated from García Criado et al (2020). Observed changes in shrub dynamics were based on field data (e.g. ecological monitoring, dendroecology), high-resolution remote sensing data (e.g. repeat photography, aerial photos), or both. Sites with mixed shrub dynamics typically had some species or species groups that were stable along with others that increased. Note that individual studies assessed changes in shrub dynamics during different time periods that do not fully overlap with observed changes in tundra greenness derived using the Landsat satellites. Background topographic data is from NOAA. Adapted by permission from Springer Nature Customer Service Centre GmbH: Springer Nature, Nature Communications, Summer warming explains widespread but not uniform greening in the Arctic tundra biome, Berner et al, Copyright © 2020, The Author(s). CC BY 4.0.
Download figure:
Standard image High-resolution imageIn tundra ecosystems, summer NDVI tracks broad spatial patterns of plant productivity (Boelman et al 2003, Street et al 2007, Kushida et al 2015, Sweet et al 2015, Berner et al 2020) and aboveground biomass (Boelman et al 2003, Walker 2003, Jia et al 2006, Raynolds et al 2012, Johansen and Tømmervik 2014, Berner et al 2018). The leafy canopies of tall deciduous shrubs can strongly affect summer NDVI (Riedel et al 2005), and thus tundra with greater deciduous shrub cover (Boelman et al 2011, Blok et al 2011b, Pattison et al 2015) and aboveground biomass (Jia et al 2003, Riedel et al 2005, Kushida et al 2009, 2015, Greaves et al 2016, Berner et al 2018) tends to have higher summer NDVI. Nevertheless, aboveground biomass corresponded much more strongly with drone-derived canopy height than NDVI in a shrub tundra landscape. These results indicate that shrub dynamics may not entirely be captured by NDVI records at landscape scales (Cunliffe et al 2020). Moreover, some tundra systems (e.g. wetlands) can have high summer NDVI despite few, if any, shrubs (Bartsch et al 2020), and relationships between summer NDVI and tundra biophysical characteristics are typically non-linear and scale dependent (Cunliffe et al 2020, Myers-Smith et al 2020).
2.2. Links between Arctic greening and observed shrub dynamics
Satellite-observed spectral greening relates to increasing shrub cover and growth in parts of the Arctic. Spectral greening has been directly linked in several studies with increasing shrub cover mapped using repeat high-resolution aerial photos and satellite imagery. For example, Landsat NDVI strongly increased from 1984 to 2014 in Siberian alder (Alnus viridis) patches that established since the 1960s across five tundra landscapes in northwestern Siberia (Frost et al 2014). Similarly, Landsat NDVI increased from 1985 to 2011 widely across a study area in the Western Canadian Arctic and was linked to increasing cover of mountain alder (Alnus crispa) and dwarf birch (Betula nana and B. glandulosa) (Fraser et al 2014). Historical reliance on coarse-resolution AVHRR NDVI datasets for spectral greening analyses has hindered direct comparisons with shrub expansion mapped with high-resolution aerial photos or satellite imagery. However, moderate- and high-resolution satellite NDVI time series are increasingly being used to assess Arctic tundra vegetation productivity changes (e.g. Arndt et al 2019, Berner et al 2020) and hold considerable promise to further elucidate how changes in shrub cover contribute to spectral greening in the Arctic.
Satellite-derived summer NDVI time series have also been linked with interannual variability in shrub growth at locations across the Arctic. For instance, summer NDVI time series have been shown to covary with annual shoot elongation of an evergreen shrub, Arctic bell-heather (Cassiope tetragona), and with annual radial growth of three deciduous shrub genera that are widespread in the Low Arctic: willow (Salix spp.), alder (Alnus spp.), and birch (Betula spp.) (e.g. Weijers et al 2018b, Andreu-Hayles et al 2020, Berner et al 2020). AVHRR NDVI time series strongly correlated with annual growth of Woolly willow (Salix lanata) from 1981 to 2005 across a tundra landscape in northwestern Russia (Forbes et al 2010). Subsequent studies linked AVHRR NDVI time series with annual shrub growth across other tundra landscapes in Russia (Blok et al 2011a, Macias-Fauria et al 2012), Alaska (Andreu-Hayles et al 2020), and Canada (Ropars et al 2015, Weijers et al 2018a). Comparisons between Landsat NDVImax and 22 shrub growth chronologies synthesized from six Arctic countries revealed moderate correlations (median Spearman correlation (rs) = 0.42) (Berner et al 2020). Several studies also documented positive trends in annual shrub growth concurrent with remotely sensed spectral greening trends (Forbes et al 2010, Ropars et al 2015, Andreu-Hayles et al 2020).
Nevertheless, neither AVHRR, MODIS, nor Landsat NDVI correspond with shrub growth in all tundra landscapes (Blok et al 2011a, Andreu-Hayles et al 2020, Berner et al 2020, Myers-Smith et al 2020), potentially because shrubs are but one component of varying dominance in plant communities that are typically intermixed within a mosaic of land cover types (Forbes et al 2010, Myers-Smith et al 2020). It also remains unclear how shrub radial growth or shoot elongation relates to changes in shrub leaf area, biomass, or landscape productivity (Andreu-Hayles et al 2020, Myers-Smith et al 2020). As a result, covariation between remotely sensed NDVI and shrub growth suggests increasing shrub growth could contribute to spectral greening, but the overall contribution of increasing shrub growth to greening remains uncertain relative to contributions from other PFTs in the Arctic.
2.3. Evidence for increasing shrub cover, growth, and biomass in the Arctic
Tundra shrubs have undergone conspicuous increases in cover, abundance, height, and growth concurrent with warming trends during the past decades in parts of the Arctic (Tape et al 2006, Rundqvist et al 2011, Myers-Smith et al 2011b, Normand et al 2013, Frost and Epstein 2014, Andreu-Hayles et al 2020, García Criado et al 2020). Species undergoing these increases include birch (Betula spp.), alder (Alnus spp.) and willow (Salix spp.) (Myers-Smith et al 2011a, Lantz et al 2013, Frost and Epstein 2014, Andruko et al 2020), dwarf evergreen shrubs including Arctic bell-heather (C. tetragona) and cowberry (Vaccinium vitis-idaea), and semi-deciduous shrubs such as Arctic avens (Dryas integrifolia) (Wilson and Nilsson 2009, Vowles et al 2017). Shrubification occurs through infilling of existing patches, increasing growth, and advancing shrublines (Myers-Smith et al 2011a).
These ecological changes have been documented in many parts of the Arctic (figure 2) through ecological monitoring (Rundqvist et al 2011), dendroecology (Boulanger-Lapointe et al 2014, Andreu-Hayles et al 2020), repeat oblique photography (Tape et al 2006, Lantz et al 2013), and high-resolution airborne and satellite remote sensing (Frost et al 2013, Moffat et al 2016). Site-level studies have reported substantial increases in shrub cover across Arctic sites, including Alaska, USA (Hollister et al 2005), northern Canada (Hill and Henry 2011), Greenland (Callaghan et al 2011), and Sweden (Becher et al 2018), among many others. While there have been many studies that reported recent shrub expansion, we note that there may be publication bias in reporting findings that exhibit changes in shrub cover. For instance, stable or decreasing shrub cover or growth has been found using ecological monitoring at many sites in the International Tundra Experiment dataset (Elmendorf et al 2012a, Bjorkman et al 2018) and reported in sites in northern Alaska and southeast Greenland (Daniëls and de Molenaar 2011, Villarreal et al 2012), among many others.
Repeat oblique photography has revealed tundra shrubification in Alaska (Sturm et al 2001b , Tape et al 2006, 2012, Brodie et al 2019), western Canada (Danby et al 2011, Mackay and Burn 2011, Moffat et al 2016), eastern Canada (Fraser et al 2011, Tremblay et al 2012), and southwest Greenland (Jørgensen et al 2013). Most of these studies relied on ground-based oblique photographs. Tape et al (2006), from an analysis of 202 pairs of oblique aerial photographs collected between ∼1950 and ∼2000, found widespread expansion of alder, willow, and dwarf birch along hillslopes and valley bottoms on the Alaskan North Slope.
For example, analysis of aerial photos from 1980 to 2013 showed tall shrub and dwarf shrub cover increased at 55% and 74%, respectively, of 38 study sites across the Tuktoyaktuk Coastlands in the Northwest Territories, Canada (Moffat et al 2016). On the other side of the Arctic, shrub cover change was evaluated at ten study sites spanning northern Siberia using high-resolution photographs from Cold-war era spy satellites (1965–1969) and recent (2009–2011) high-resolution commercial satellite imagery (Frost and Epstein 2014, Frost et al 2014). Shrub cover exhibited little net change (−0.8%) at one study site but increased 5%–26% (mean = 13%) across the other nine study sites, particularly in landscape positions with active disturbance regimes (e.g. permafrost-related patterned-ground, floodplains, hillslopes) (Frost and Epstein 2014). While attention is often paid to areas with shrub expansion, aerial photos also reveal areas with little to no change in shrub cover during recent decades (e.g. Plante et al 2014, Jorgenson et al 2018). Overall, high-resolution remote sensing analyses illustrate extensive increases in shrub cover during the last four to seven decades in the Arctic, while also underscoring that change did not uniformly occur across tundra landscapes.
3. Environmental and climatic drivers, interactions, and mechanisms of shrub expansion
3.1. Climate warming
Warming temperatures have been reported at biome-wide scales across the tundra (IPCC 2013, AMAP 2017) and are associated with increasing shrub cover across the Arctic (Myers-Smith et al 2015). While Arctic plant communities are generally sensitive to warming, the responses are site-dependent and heterogeneous (Hollister et al 2005a, Bjorkman et al 2020, García Criado et al 2020, Myers-Smith et al 2020). Shrub species differ substantially in their responses to climate change because of a variety of factors other than warming (e.g. site conditions, soil moisture, snow-dynamics, plant-specific responses) (García Criado et al 2020, Myers-Smith et al 2020). Patterns of plant community responses to warming are consistent between monitoring and experimental warming methods, although space-for-time approaches do not appear to be appropriate for quantifying the rate or magnitude of change (Elmendorf et al 2012a, 2012b, 2015) and short- and long-term responses are expected to differ (Bouskill et al 2020). The most apparent link between climate and shrub expansion is the correlation of temperature with shrub growth, abundance, and recruitment. Observational studies, based on remote or ground surveys over time or space and on warming experiments, have found higher shrub growth and recruitment with warmer temperatures. The trend of warmer summers correlates with shrub expansion across the Arctic tundra (Myers-Smith and Hik 2018, Weijers et al 2018b, Berner et al 2020).
The response of shrub growth to warming is spatially heterogeneous, however, with higher temperature sensitivity in the European Arctic than in North America and at sites with greater soil moisture and taller shrubs (Myers-Smith et al 2015). Plant and ring width growth that correlate with summer NDVI (section 2.2) have also been found to correlate with summer temperatures (Myers-Smith et al 2015). Annual growth of alder and willows have been found to be climate sensitive around the circumpolar Arctic (Myers-Smith et al 2015) including in Arctic Alaska (Tape et al 2012, Andreu-Hayles et al 2020), northwest Russia (Forbes et al 2010), Arctic Canada (Boulanger-Lapointe et al 2014, Myers-Smith and Hik 2018, Weijers et al 2018a), and Greenland, Norway and Svalbard (Jørgensen et al 2015, Weijers et al 2018a). These trends are corroborated by large-scale dendroecological syntheses (Myers-Smith et al 2015).
Temperature manipulation experiments allow more controlled investigation of warming impacts on shrub productivity. Meta-analyses and most experiments report that warming promotes growth (Walker et al 2006) and germination (i.e. seed biomass, cumulative germination, germination rate, peak germination (Klady et al 2011)) unless moisture or other conditions are limiting. Experimental warming can also increase seedling mortality, resulting in no net effect on establishment (Milbau et al 2017). Biome-wide remote sensing and modeling also confirm that warming is leading to more favorable conditions for shrubs. For example, the extent of Arctic areas where vegetation is limited by temperature declined over the past three decades (Keenan and Riley 2018). Although warming tends to expand shrub ranges into currently colder locations, it can also cause contraction at the warmer or southern edge of current shrub ranges (Bokhorst et al 2018), due to population-level variation in temperature optima (Kueppers et al 2017) or due to treeline advance, as seen between 1900 and 2008 (Harsch et al 2009).
3.2. Soil moisture and snow dynamics
Soil moisture, snow dynamics, and other climate-related factors can also influence shrub growth and establishment (Martin et al 2017). Furthermore, air temperature often covaries with soil moisture, snow dynamics, active layer depth, nutrient availability, and other environmental conditions that affect shrub success (figure 1). These co-varying factors are difficult to control in observational studies and receive less assessment compared to air temperature (Martin et al 2017, Myers-Smith et al 2019a). When these factors have been investigated, their interactions with the warming effect were variable. Multiple studies have reported greater shrub expansion in more moist sites. For example, Naito and Cairns (2011) found that shrubs expand more into areas with higher topographic wetness index and closer to the riverbank in the North Slope of Alaska. Boulanger-Lapointe et al (2014) found willow cover and seedling density was high in sites with elevated soil moisture but limited by water availability in dry sites in the High Arctic of Greenland and Canada. García Criado et al (2020) found that tundra woody cover changed more rapidly in wetter sites. Higher temperature sensitivity of shrub growth was found for wetter vs drier sites (Myers-Smith et al 2015, Ackerman et al 2017). Field surveys and warming experiments indicate that moisture limitation can reduce shrub growth, recruitment, and abundance (Elmendorf et al 2012a, Myers-Smith et al 2015, Li et al 2016, Ackerman et al 2017). Across all of these studies, the availability of soil moisture strongly interacts with warming. Soil moisture also plays a critical role in determining the trajectory of vegetation under warming (Elmendorf et al 2012b, Ackerman et al 2017, Bjorkman et al 2018). Bjorkman et al (2018), in a large-scale analysis of the relationships between plant traits, warming, and soil moisture, highlighted the importance of soil moisture and concluded that the trajectory of changes in plant traits and ecosystem function under future warming would depend on soil moisture.
As glaciers retreat and permanent snow cover shrinks, reduced melt-water supply in the growing season might induce moisture limitation in extended regions (Boulanger-Lapointe et al 2014). Warming can lead to earlier snowmelt, which can affect soil moisture and lengthen the growing season, which have been found to promote shrub growth in some studies (Hill and Henry 2011, Wilcox et al 2019). However, because snowpack protects shrub shoots and seedlings from damage caused by fungal and insect attacks and frost, earlier snowmelt due to warming has also been found to offset some positive impacts of warming (Bokhorst et al 2009, Wheeler et al 2016).
3.3. Topography
Topography is an important distal controller of vegetation growth in many ecosystems. The more proximal vegetation growth controllers influenced by topography include soil moisture, snowpack redistributions, lateral nutrient and oxygen fluxes, relatively static soil properties (e.g. texture (perhaps affected by erosion), depth), soil redox and nutrient states, snow cover and properties, disturbance, and light (figure 1). Although these factors affect all ecosystems, the conditions in tundra systems (e.g. permafrost, patterned ground, thermokarst) lead to unique controls on shrub growth. The three observed categories of recent shrub increases (i.e. infilling, growth increases, and range spread) identified by Myers-Smith et al (2011a) will each be affected by different combinations of these mechanisms. However, there are relatively fewer observational studies that quantify the relative importance of these mechanistic controls than in, e.g. temperate forests.
Observations of relationships between tundra shrub cover change and topographically driven processes have been used to infer mechanisms affecting these interactions. Many studies (Chapin et al 1988, Epstein et al 2004, Naito and Cairns 2011, Myers-Smith et al 2015, Lara et al 2018, Campbell et al 2020) indicate that tundra plant productivity is strongly affected by topographically driven hydrology. Additional observed factors associated with topography that affect shrub growth include snow properties (Boulanger-Lapointe et al 2016), thermokarst in patterned ground (Frost et al 2013, Huebner and Bret-Harte 2019), soil properties, and cryoturbation (Ropars and Boudreau 2012, Frost et al 2014, Swanson 2015).
A number of land models have been applied to analyze tundra shrub dynamics (Epstein et al 2000, Euskirchen et al 2009, Lawrence and Swenson 2011, Bonfils et al 2012, Miller and Smith 2012, Zhang et al 2013, Druel et al 2019), but none of these models explicitly represent topographical variation at the relevant spatial scales. We identified only a few studies that have applied land models resolving topographic gradients that evaluated the effects on vegetation. Mekonnen et al (2021b) showed that hillslope topography and thereby hydrology strongly controlled historical and future shrub growth. At the hill crest, canopy water stress and low plant nitrogen uptake led to low modeled shrub biomass. In the mid-slope position, intermediate soil water content reduced shrub water and nitrogen stress, leading to higher shrub biomass. In the lower-slope position, saturated soil conditions reduced soil oxygen concentrations, nutrient availability and uptake, and plant biomass. An analysis with simulations that ignored topographical gradients and gridcell inter-connectivity underestimated mean shrub biomass and over- or under-estimated shrub productivity at the various hillslope positions.
The emergent patterns of shrub expansion responses to climate warming and changes in soil moisture summarized above interact with a range of processes, including permafrost thaw, nutrient cycling, and disturbance, as discussed in the following sections.
3.4. Permafrost thaw
Thawing of permafrost (Brown and Romanovsky 2008, Schuur et al 2015, Hugelius et al 2020, Mekonnen et al 2021a) and thermokarst development (Schuur et al 2007, Jones et al 2015) may alter soil moisture and thermal regimes and thus affect nutrient availability. Evidence from many Arctic sites suggests that a deeper active layer promotes shrub expansion (Martin et al 2017). Recent and projected warming are expected to deepen the active layer heterogeneously depending on climate and soil properties, and thereby increase nutrient availability (Mekonnen et al 2018b). These environmental conditions, though connected to air temperature through ecosystem and climate feedbacks, directly influence shrub growth and establishment.
Thawing of ice-rich permafrost or the melting of massive ice may lead to a landscape deformation process resulting in thermokarst development (van Everdingen 2005). Alterations of the ground surface, such as ground temperature, thaw depth, and soil moisture could provide favorable conditions for shrub growth (Schuur et al 2007, Lantz et al 2010, Frost et al 2013). Thermokarst disturbances of tundra surfaces will increase access to bare soils initially covered by a thick organic mat and could increase germination of shrub species (Lloyd et al 2003, Lantz 2017, Mikhailov 2020). Observational studies have reported increased soil temperature, thaw depth, nutrient availability, and snowpack in sites associated with thermokarst subsidence and landform deformation, and concurrent increases in shrub growth (Lloyd et al 2003), but the effects vary with thermokarst type and process (supplementary material). In thermokarst depressions on tundra hills with harsher conditions, protection from wind and frost burn may play essential roles in shrub expansion (Lantz 2017).
3.5. Nutrient cycling
As high-latitude soils warm, availability of plant nutrients (i.e. nitrogen and phosphorus) is expected to increase (figure 3). This increase stems from kinetically controlled mineralization rates, which are strong functions of temperature (Nadelhoffer et al 1991, Blok et al 2018), and deepening active-layer exposing previously frozen organic matter (Salmon et al 2018) for decomposition, nutrient mineralization, and plant assimilation (Keuper et al 2017, Blume-Werry et al 2019, Hewitt et al 2020). Furthermore, observations of increasing wintertime respiration, particularly under deeper snowpack, could play an important role in releasing nutrients over the 21st century (Schimel et al 2004, Natali et al 2019). However, whether nutrients released during subnivean activity are assimilated by plants or lost hydrologically during snowmelt remains an open question (Grogan and Jonasson 2003, Edwards et al 2006, Koven et al 2015, Riley et al 2018).
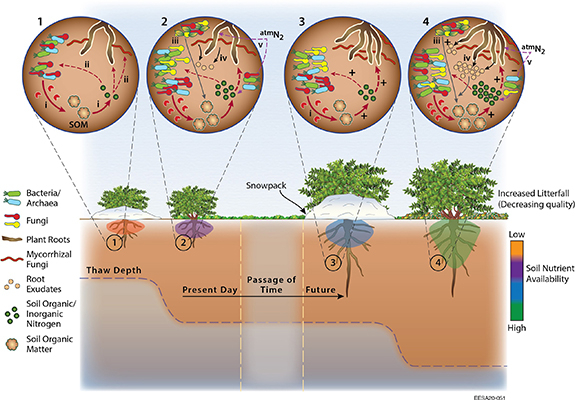
{kind=link}
{kind=link}
Figure 3. The schematic depicts the changes to belowground ecosystems and nutrient cycling potentially occurring under increased shrub growth and cover. Each circular panel represents the soil environment underneath the shrubs during different periods of the year: under wintertime snowpack, and during summer. Broadly, from present day to a future date the figure shows increased shrub height, cover, nutrient cycling, and availability, as described in the text. (i) Exoenzyme production hydrolyzes complex soil organic matter releasing simple organic compounds (monomers) and nutrients (organic and inorganic) that are (ii) available for rapid assimilation by shrub roots, associated mycorrhizae, and microorganisms. As winter transitions to summer, plant activity increases, playing a critical role in belowground nutrient cycling: plant litterfall (iii) can be decomposed by soil fauna, yielding soil organic matter (SOM), and contributing to the cycling of soil nutrients. Furthermore, plant exudation (iv) can stimulate the microbial community, and encourage mining of existing SOM to yield inorganic nitrogen and phosphorus. Free-living and symbiotic nitrogen-fixing bacteria (v) can further increase soil nitrogen stocks through fixation of atmospheric nitrogen. Under future climate scenarios, elevated shrub growth, deeper rooting depths, and deeper snowpacks are predicted to increase subsurface wintertime activity, leading to higher nutrient availability for assimilation by plants and microbes, and increased wintertime respiration. Transitioning to summertime leads to a higher litterfall associated with a larger and denser canopy. The litter associated with shrubs has a higher C:N ratio, which increases the fungal to bacterial ratios, as conditions encourage the growth and colonization of fungi. However, the fate of diazotrophic microorganisms over time is generally uncertain. For example, an increase in soil nitrogen availability through other mechanisms could reduce the requirement for atmospheric nitrogen fixation. Signs (±) on soil panels 3 and 4 represent increases or decreases in processes over time, relative to the corresponding panels 1 and 2.
Download figure:
Standard image High-resolution image{kind=link}
How higher nutrient availability shapes vegetation composition and productivity has been examined directly through nutrient manipulation experiments (Mack et al 2004, DeMarco et al 2014b, Prager et al 2020) and indirectly through long-term observations across spatial gradients (Pelletier et al 2019). In general, increased nutrient availability enhances shrub productivity, coverage, and biomass (Shaver and Chapin 1980, Mack et al 2004), which can lead to taller shrubs with higher leaf nitrogen content (Bjorkman et al 2018, Prager et al 2020). A feedback loop may subsequently emerge (figure 3) whereby increased shrub biomass and height under warmer temperatures and elevated nutrient availability can lead to a deepening snowpack, insulating the under-snow soil, and further increasing nutrient mineralization rates and nutrient availability into the summer (Chapin et al 2005, Sturm et al 2005b, Bjorkman et al 2018, Hicks et al 2020). However, interactions between snow depth and belowground activity are complex. A controlled manipulation study showed no impact on nutrient cycling of a deepening snowpack (Myers-Smith and Hik 2013), while litter quantity and quality were likely more significant factors driving decomposition (DeMarco et al 2014a).
Traits that control shrub-ectomycorrhizal (ECM) associations may also provide mechanistic insight into the competitive dynamics of tundra vegetation. For instance, mycorrhizal networks exist in tundra and facilitate belowground inter-plant carbon transfers, and thus may alter competitive abilities (Deslippe et al 2011, Deslippe and Simard 2011). Tundra shrubs, and associated ECM fungi, have been shown to have higher maximum uptake rates (VMAX) than graminoids (Zhu et al 2016), and allocate significant resources towards fine root biomass and therefore nutrient uptake capacity (Vamerali et al 2003, Iversen et al 2015). Such competitiveness for nutrients could explain consistent trait responses across nutrient-manipulation studies. For example, in a recent study examining plant traits under nutrient fertilization, Prager et al (2020) found that only deciduous shrubs had significantly greater leaf N at high levels of nutrient addition. Furthermore, under similar nutrient-enrichment conditions, deciduous shrubs, but not graminoids, increased foliar N (Heskel et al 2012). Similarly, Bret-Harte et al (2001) noted that elevated nutrient availability increased shrub leaf area index and canopy density, resulting in light limitation of understory species. These relationships imply that high foliar nutrient levels and the ability to acquire soil nutrients are beneficial traits in high-latitude environments. However, Bjorkman et al (2018) noted that traits such as leaf nitrogen content generally decreased with warming in dry sites, while increasing in wetter conditions in analyses at the community-level.
Finally, the importance of post-growing season nutrient uptake has become increasingly clear in recent years (Riley et al 2018). Belowground plant activity (e.g. root growth) continues after the cessation of photosynthetic activity (Iversen et al 2015, Blume-Werry et al 2016), while nutrient uptake continues into the winter (Chapin and Bloom 1976, Andresen and Michelsen 2005). Shrubs have been noted to be particularly active during spring, relative to other tundra plants (Weih 2000, Larsen et al 2012), which could lead shifts in phenology under a warmer climate (Mekonnen et al 2018b, Oberbauer et al 2013).
3.6. Wildfire
Wildfires can burn and remove shrubs, and thus reduce post-fire shrub populations in the short-term. Wildfires often alter soil temperature (Jiang et al 2015), surface litter and soil organic carbon stocks (Grosse et al 2011, Mack et al 2011, Chen et al 2021), and seedbed quality, thus affecting regeneration and shrub establishment (Lantz et al 2010, Bret-Harte et al 2013). Removal of an insulating surface litter layer increases the active layer depth (Iwahana et al 2016, Michaelides et al 2019). These factors may alter soil organic carbon stocks, soil moisture, and nutrient dynamics and thus affect post-fire competition and successional trajectories in the tundra. Fire may also impact shrub performance by altering mycorrhizal symbionts. Hewitt et al (2013) concluded that the resprouting strategy of tundra shrubs makes the dominant mycorrhizal fungi more resilient to fire by maintaining an inoculum source on the landscape after fire. As a result, resprouting shrubs may facilitate post-fire vegetation regeneration and potentially shrub expansion under future warming and fire regimes.
Wildfires increase shrub expansion in tundra regions over multi-decadal timescales, although short-term observations of disturbed tundra indicate negative influences of wildfires on shrubs. Based on paleoecological studies (e.g. Higuera et al 2008, Hu et al 2010) and model simulations (Rupp et al 2000, Mekonnen et al 2019, Bouskill et al 2020), the predicted frequent and larger wildfires in the Arctic under future climate are expected to increase Arctic shrub expansion. Several studies across the tundra reported increases in shrub growth and distribution in older burn scars, although post-fire shrub recovery may last for more than ten years. For instance, graminoid biomass was shown to recover four years after the Anaktuvuk River Fire (ARF) on the North Slope, Alaska (Bret-Harte et al 2013). However, shrubs did not recover to pre-fire conditions (Jandt et al 2012, Bret-Harte et al 2013) or were recovering slowly by the fourth year after the ARF (Jandt et al 2012). Narita et al (2015) and Iwahana et al (2016) observed vegetation and thaw depth changes five to ten years after a 2002 tundra fire on the Seward Peninsula, Alaska. They found evergreen shrub cover was still substantially lower five years after the fire and had not recovered ten years after the fire. In contrast, graminoid and deciduous shrub cover had increased over the same period. The long-term increase in shrubs following fire was also shown in other studies on the Seward Peninsula, Alaska and in Western Siberia (Racine et al 2004, Heim et al 2019). These studies show that fire promotes shrub growth and expansion and thus alters ecosystem carbon balance, although fire may reduce shrub growth in the short term.
Short-term increases in active layer depth are a common feature following tundra wildfires. Depending on site conditions, post-fire thaw depth may recover to the level of unburned sites in about ten years (Iwahana et al 2016), or persist longer (Rocha et al 2012), while increased shrub growth and expansion may continue for decades. Spatial variation of active layer depth may also be related to plant community composition, with deeper thaw corresponding to graminoid-rich areas and shallower thaw corresponding to shrub-rich areas (Narita et al 2015). The active layer depth returned to pre-fire levels ten years after the 2002 Kougarok fire, Seward Peninsula, Alaska (Narita et al 2015, Iwahana et al 2016). Active layer was deepest (52.3 cm) 10–11 years following a fire and gradually returned to unburned levels thereafter.
Some areas burned in recent Kougarok fires experienced thermokarst, especially polygonal depressions along lines of ice-wedges (Iwahana et al 2016, Tsuyuzaki et al 2018). Frost et al (2020) studied vegetation changes after 1971–1972, 1985, 2006–2007, and 2015 tundra fires on the Yukon-Kuskokwim Delta, Alaska. Shrub cover was lower in younger fire scars (one to three years) than adjacent unburned areas, but higher shrub cover occurred at sites with older fires (10–46 years). Using radiocarbon dating, aerial photography, and climate proxy data, Jones et al (2013) identified tundra fires that likely occurred between AD 1880 and 1920 on the North Slope, Alaska. They found degradation of ice-rich permafrost and increased shrub vegetation with taller canopy height than surrounding unburned areas. In the Mackenzie Delta Uplands, Lantz et al (2013) also found the highest shrub coverages (92%–99%) in old-burned (about 40 years) tundra among their studied tundra areas. These results suggest changes in recovery times of thaw depth and shrub biomass following fire may subsequently alter post-fire successional trajectories and ecosystem carbon balance.
3.7. Herbivory and grazing
Herbivory is an important factor that affects shrub growth in the tundra (Olofsson et al 2004, Post and Pedersen 2008, Tape 2011). Several exclosure experiments have demonstrated that herbivory may alter vegetation composition (Pajunen et al 2008, Ravolainen et al 2011) and reduce climate-driven shrub expansion (Olofsson et al 2009). For instance, in a ten year field experiment with permanent plots with treatments of reindeer only vs all mammalian (small mammals and reindeer) exclosures at four forest–tundra ecotone locations in northern Fennoscandia, shrub abundance was generally shown to increase with herbivore exclusions (Olofsson et al 2009). In a three year exclosure experiment in a Low Arctic site in Norway, the biomass of forbs, deciduous shrubs, and herbaceous plants were shown to increase by 40%–50% in the absence of herbivory (Ravolainen et al 2011). The effects on different tundra PFTs may also vary with herbivores. For instance, rodents may prefer mosses and dwarf shrubs (Moen et al 1993, Dahlgren et al 2007), while reindeer and wild caribou were shown to prefer deciduous shrubs and lichens (Herder et al 2003, Post and Pedersen 2008). Herbivores can also affect seedbed quality, seedling establishment, and growth of tundra plants (Munier et al 2010). These processes can strongly alter tundra PFT prevalence, suggesting that herbivore abundance and distribution may have a direct impact on shrub expansion across the tundra.
Climate warming may have direct and indirect effects on plant–herbivore interactions (Olofsson et al 2009) and on changes in the abundance of herbivores and their predators (Ims and Fuglei 2005). Shrub expansion and shifts in vegetation composition driven by climate warming can alter the composition and quality of forage for herbivores (Kitti et al 2006, Doiron et al 2014). Increases in shrub cover may also increase snowpack height that may affect timing of snowmelt, growing season length, and forage access to herbivores (Berg et al 2008). Indirectly, climate warming may also alter predator–prey interactions, thus altering herbivore populations (Legagneux et al 2014) and their impact on shrubs.
3.8. Plant traits and competition
Warmer climate, enhanced nutrient cycling (which will lead to increased nutrient availability), and disturbance modify competitive interactions of tundra plants and thereby may result in changes in relative shrub abundance (Mack et al 2004, DeMarco et al 2014b, Zamin et al 2014, Prager et al 2020). Competitive interactions among PFTs, as mediated through functional traits, strongly affect community assembly through competition for light, water, and nutrients under changing climate (Soudzilovskaia et al 2013, Myers-Smith et al 2019b). Several plant traits (e.g. plant height, leaf nutrients and optical properties, phenology, morphology, root traits, and axial hydraulic resistance) are known to differ among tundra PFTs (Chapin et al 1996a, Iversen et al 2015, Bjorkman et al 2018, Myers-Smith et al 2019b, Thomas et al 2019). As a result, tundra PFTs differ in their abilities to acquire and retain resources. Across ecosystems, carbon and nutrient investment and retention strategies in leaves, stems, and roots partly explain PFTs' competitive abilities under changing climate (Chapin et al 1996a, Westoby et al 2002, Wright et al 2004, Soudzilovskaia et al 2013). Differences in traits may also affect emergent PFT variation in phenology, irradiance, CO2 fixation rate, and water uptake and thereby each PFT's competitive growth. Structural traits such as plant height respond strongly to changes in growing conditions in tundra ecosystems (Bjorkman et al 2018), yet do not differ strongly among PFTs in tundra plants (Thomas et al 2019).
With increases in nutrient availability, such as those expected over the 21st century (Mekonnen et al 2018b), shrubs may grow faster leading to greater carbon gains per N investment, resulting in higher woody carbon stocks (Sistla et al 2013), with longer turnover times and higher plant carbon to nitrogen (C:N) ratios (Weintraub and Schimel 2005). Shrubs associated with symbiotic N2 fixation (Densmore 2005, Salmon et al 2019) may also compete more effectively due to their independent supply of N.
The rapid growth of shrubs with greater height and leaf area (e.g. Hudson et al 2011, Elmendorf et al 2012a) have led to the competitive exclusion of shade-intolerant species, such as lichens and mosses (Cornelissen et al 2001, Walker et al 2006, Pajunen et al 2011, Elmendorf et al 2012a, Fraser et al 2014) at some warming experiment sites, further increasing shrubs' ability to compete. While exclusion via light has not been observed at all sites (e.g. Elmendorf et al 2012a), light competition can impact plant functional diversity and community structure. Plant etiolation responses to light attenuation is an important trait that controls their ability to effectively compete under shading (Havström et al 1993, Chapin et al 1996b). Thus, traits that control maximum canopy height and greater carbon uptake may result in shrubs being more competitive than other PFTs in tundra ecosystems in a warmer climate (Mekonnen et al 2018b).
4. Impacts of shrub expansion on ecosystem carbon balance
Recent changes in vegetation composition, particularly shrub expansion, may alter the tundra ecosystem carbon balance directly through effects on (a) ecosystem net primary productivity and thus biomass and (b) surface litter inputs, and thus soil organic carbon and respiration. Shrub expansion can also indirectly affect ecosystem carbon cycling through (a) snow–shrub interactions (Liston et al 2002, Sturm et al 2005a, Myers-Smith and Hik 2013, Marsh et al 2010), (b) permafrost degradation (Blok et al 2010, Lawrence and Swenson 2011, Nauta et al 2015), (c) surface energy balance (Chapin et al 2000, Bonfils et al 2012, Lafleur and Humphreys 2018), (d) nutrient cycling (DeMarco et al 2014a, Christiansen et al 2018a, Wang et al 2018, Prager et al 2020), and (e) ecosystem carbon turnover time (Parker et al 2015, Ravn et al 2020). The overall impacts of shrub expansion on ecosystem carbon balance depend on complex interacting and rapidly changing climatic and environmental factors.
4.1. Ecosystem productivity and carbon balance
Several studies reported increases in plant carbon uptake inferred from Arctic greening and increases in shrub biomass (section 2). Circum-Arctic trends inferred based on multi-decadal changes in remote sensing observations (Rouse et al 1974) indicate increases in plant productivity (Tucker et al 2001, Olthof et al 2008, Verbyla 2008) and shrub growth and biomass (Frost et al 2013, Moffat et al 2016) across much of the Arctic tundra biome during the past decades (sections 2.2 and 2.3; figure 2). Observed increases in carbon uptake and thus shrub growth and biomass were also shown based on a systematic literature review updated from García Criado et al (2020) (figure 2). These results have been supported with a shrub ring width chronology analysis (Blok et al 2011b) that showed increases in shrub growth. Dendroecological measurements indicate changes in shrub radial growth and establishment at sites across the Arctic (Myers-Smith et al 2015). Shrub radial growth has been found to commonly relate to above-ground biomass Moullec et al 2019 and remotely sensed NDVI (Forbes et al 2010, Ropars et al 2015, Andreu-Hayles et al 2020, Berner et al 2020, Myers-Smith et al 2020), though not always (Weijers et al 2018b).
Further, results from field and remote sensing observations were corroborated by several natural and artificial warming experiments. Most of these experiments have shown an increase in shrub net primary production. For instance, long-term plot observations from 46 sites across the tundra (Elmendorf et al 2012b) showed overall increases in height and abundance of shrubs, although responses varied with site conditions (Myers-Smith et al 2015). In other warming experiments across the tundra, increases in height and biomass of tall deciduous shrubs were shown (van Wijk et al 2004, Walker et al 2006, Zamin and Grogan 2012, Sistla et al 2013). Gains in net primary productivity that led to shrub growth were primarily driven by enhanced CO2 fixation via higher N mineralization, deeper thaw depth, and thus increased nutrient availability (Campioli et al 2013, DeMarco et al 2014b). Nutrient availability and plant productivity may further be enhanced by biological N2 fixation through symbiotic associations with bacteria (e.g. Alder), and primed by increased root carbon allocation (Rhoades et al 2001, Densmore 2005). This robust range of observations and warming experiments suggest enhanced plant carbon uptake that led to increased biomass of shrubs in response to warming.
Expansion of shrubs also increases carbon losses through ecosystem respiration (Parker et al 2015, Phillips and Wurzburger 2019) by increasing litter inputs (Liston et al 2002, Myers-Smith and Hik 2013, Christiansen et al 2018b, Kropp et al 2018) and active layer depths (Blok et al 2010, Frost et al 2018, Wilcox et al 2019). The net ecosystem carbon balance in a shrub-dominated site depends on contrasting responses of ecosystem carbon uptake vs respiration. Several meta-analyses of long-term ecosystem warming experiments (Arft et al 1999, Dormann and Woodin 2002, Rustad et al 2001, Walker et al 2006, Elmendorf et al 2012a) have shown that ecosystem responses to warming are spatially heterogeneous and dependent on the climate zone, site conditions (e.g. local topography, soil properties, surface and subsurface hydrology), and PFT composition (via litter inputs). Since woody shrubs have the highest C:N ratio among tundra PFTs and their woody litter decomposes relatively slowly, their relative increase across the Arctic may enhance ecosystem carbon storage (Weintraub and Schimel 2005, Heskel et al 2013). However, increases in net carbon uptake from shrub expansion may be offset by concurrent increases in ecosystem respiration (Biasi et al 2008). In-situ measurements in shrub-dominated sites indicate contrasting responses of shrub biomass and soil organic carbon. Shrub expansion and the subsequent increase in biomass (Berner et al 2018, García Criado et al 2020) and litter inputs (Elmendorf et al 2012b) may alter decomposition rates of soil organic carbon (Myers-Smith and Hik 2013, Sistla et al 2013, Parker et al 2015, Lynch et al 2018, Gagnon et al 2019, Christiansen et al 2018a). Flux data from 21 sites across the Arctic and boreal ecosystems have shown a strong ecosystem carbon sink for sites dominated by shrubs vs herbaceous plants (Cahoon et al 2012). However, sites with greater summer soil temperatures were shown to be carbon sources. These results suggest that shrub expansion impacts on net ecosystem exchange is site specific, and dependent on changes in biomass vs soil organic carbon stocks. We note that, although several studies in the literature reported effects of shrub expansion on biomass and decomposition of soil organic carbon (e.g. Christiansen et al 2018a, Gagnon et al 2019, Lynch et al 2018), the effects on net biome productivity have not been widely measured.
4.2. Albedo, surface energy budgets
Shrubs affect ecosystem carbon balances indirectly through effects on the surface energy balance and snowpack accumulation (Liston et al 2002, Marsh et al 2010, Nowinski et al 2010, Myers-Smith and Hik 2013). Increases in tall shrubs that grow above the snowpack reduce albedo, altering the energy balance and thus snowmelt timing (Marsh et al 2010). Sturm et al (2005a) showed from measurements at five tundra sites in Alaska that sites dominated by tall shrubs resulted in a 30% reduction in winter time albedo, compared to sites with dwarf shrubs underneath the snowpack. Increases in canopy net radiation from reduced albedo by tall shrubs may alter seasonal carbon uptake (Livensperger et al 2016, Lafleur and Humphreys 2018). Changes in spring albedo may also affect snowmelt timing (Sturm et al 2005a, Marsh et al 2010), thus resulting in earlier leaf-out and carbon uptake (Bonfils et al 2012, Livensperger et al 2016). While earlier snow-melt results in more spring snow-free days and greening (Livensperger et al 2016), it may also reduce dwarf-shrub growth, likely related to adverse effects of temperature on the early growing season (Wheeler et al 2016). Thus, reduced albedo that leads to earlier snowmelt may have a contrasting impact on spring carbon uptake.
4.3. Snow–shrub interactions
Tall deciduous shrubs can accumulate snow redistributed by wind across a landscape (Liston et al 2002, Pomeroy et al 2006). Snow fence experiments at Arctic tundra sites show that deeper snowpack promotes winter soil warming (Nobrega and Grogan 2007, Joshua Leffler and Welker 2013). Deeper snowpack insulates the soil surface, resulting in warmer soil during winter (Paradis et al 2016) and thus increased soil organic matter (SOM) decomposition (Sturm et al 2001a). Warmer soil may accelerate wintertime ecosystem respiration that may substantially contribute to non-growing season carbon loss (Natali et al 2019), but may also increase nutrient availability to plants and facilitate higher growing season biomass gains (Riley et al 2018). Deeper snow may also deepen the active layer (Nowinski et al 2010) and increase soil wetness and thus methane production (Blanc‐Betes et al 2016). While snow accumulation may enhance winter soil warming (Myers-Smith and Hik 2013) and active layer depth (Nowinski et al 2010), shrub expansion was also reported to cool soils (Myers-Smith and Hik 2013) and reduce summer permafrost thaw (Blok et al 2010) from shading of the soil surface by the greater canopy cover. These interactions may result in lower summer soil temperatures, decomposition rates, and summertime nutrient availability (Myers-Smith and Hik 2013). Thus, alteration of snowpack dynamics affects seasonal soil temperature, nutrient dynamics, and rates of ecosystem respiration.
4.4. Litter inputs to soils and decomposition
Long-term plots show increases in tall deciduous shrub growth leading to greater litter inputs (Elmendorf et al 2012b). However, increased shrub litter inputs have contrasting effects on net ecosystem carbon exchange. Shrub growth may be enhanced from greater litter inputs, which can accelerate SOM decomposition, thus increasing nutrient availability (Buckeridge et al 2010). Rapid decomposition of shrub litter may also increase SOM carbon losses through heterotrophic respiration (Nielsen et al 2019, Phillips and Wurzburger 2019). Wintertime shrub litter decomposition was shown to be accelerated from deeper snowpack that resulted in warmer soil, although spring warming, under drier conditions, was reported to reduce litter decomposition rates (Blok et al 2016). The net effect of increased shrub litter on ecosystem carbon balances depends on litter quality and quantity. For instance, Christiansen et al (2018b) reported greater litter carbon losses in tall vs low birch shrubs at Daring Lake, a mesic Arctic tundra site in Canada. Changes in litter decomposition rates and the subsequent effects on carbon uptake may alter ecosystem carbon residence times (Parker et al 2015, Ravn et al 2020).
5. Observational, theoretical, and modeling research gaps and future directions
5.1. Research gap: climate change effects on shrub expansion
We highlight two major gaps in the observational literature of climate controls on shrub expansion. First, while shrub expansion is controlled by multiple climatic and environmental conditions, most studies have focused on the direct impacts of warming or soil moisture. Yet research also suggests that warming may not be the dominant control on growth or establishment when other factors such as soil moisture, snow dynamics, permafrost thaw, nutrient cycling, and biotic activity are also considered or controlled (Martin et al 2017, Lett and Dorrepaal 2018, Myers-Smith et al 2019b). As such, there is a need for multifactorial experiments and observational analysis. Disentangling impacts of multiple factors will contribute to a better assessment of the relative influences of positive and negative feedbacks under warming, which are key to projecting future rates of shrub expansion (Myers-Smith et al 2011a, 2015). Second, most analyses are based on relatively short-term observations (<25 years) or substitute spatial patterns for longitudinal studies to quantify temporal responses. The limited time range and reliance on space-for-time approaches pose challenges in addressing time lags in shrub response to variation in environmental conditions (Büntgen et al 2015) and to adequately representing long-term responses (Elmendorf et al 2012a, Martin et al 2017, Bouskill et al 2020).
5.2. Research gap: influence of changing microbial communities on nutrient cycling
The strong interplay between shrub expansion and nutrient availability underscores a critical role for soil microbes (figure 3). Microbial communities can promote shrub expansion through, for example, the mining of nitrogen from organic compounds in response to rhizodeposition (Hicks et al 2020, Street et al 2020). However, shifts in tundra vegetation, including shrub expansion, can alter the composition and abundance of microbial functional guilds (Wallenstein et al 2007, Eskelinen et al 2009, Shi et al 2015). Shrub expansion can modify the quantity, quality, and chemical composition of SOM (McLaren et al 2017) due to increased root (Brüggemann et al 2011) and litter production (Cornelissen et al 2007), and rhizodeposition (Street et al 2020). Despite strong functional redundancy (Louca et al 2018), shifts in microbial community composition within the tundra can lead to changes in metabolic function, including, for example, increased carbohydrate utilization (Johnston et al 2019). Nonetheless, impacts on the tundra carbon cycle remain uncertain, with evidence for and against the priming of existing SOM under higher rhizodeposition and litter production (Lynch et al 2018, Hicks et al 2020, Street et al 2020).
Of particular significance to tundra carbon and nutrient cycling is the potential change in fungal–plant interactions that could emerge under shrub expansion (Clemmensen et al 2006, Bennett and Classen 2020). ECM and ericoid mycorrhizal fungi, typically partnered with deciduous and evergreen shrubs respectively (Deslippe et al 2011, Deslippe and Simard 2011, Vowles and Björk 2019, Hicks et al 2020), play an important role in the acquisition and transfer of nutrients and water to the plant (Read and Perez-Moreno 2003, Fernandez and Kennedy 2016, Hewitt et al 2020). However, the feedback to soil carbon stability remains uncertain, with evidence both for and against increased decomposition rates attributable to changes in the abundance and composition of mycorrhizal fungal (Fernandez and Kennedy 2016).
5.3. Research gap: recruitment effects on shrub expansion
By definition, shrub reproduction and establishment refer to range shifts or infilling, but controls on these processes are not well-described in the literature. It is less clear how seed germination and seedling establishment, potential bottlenecks for Arctic shrub expansion, will respond to a warmer climate (Büntgen et al 2015, Milbau et al 2017, Myers-Smith and Hik 2018). Factors controlling the recruitment of new shrub individuals need further study to predict shrubline advance and changes in PFTs driven by climate and disturbance. More studies are needed that evaluate site-specific controls on regeneration, such as seed production, suitable microsite availability, recruitment, seedling survival, and establishment (Šenfeldr and Treml 2020). The conditions that control seedling survivorship are not always the same as those that control growth of mature shrubs, yet there is little overlap among studies of these two processes (Büntgen et al 2015, Angers-Blondin et al 2018, Myers-Smith and Hik 2018). It is relatively easy to conduct manipulation experiments with seeds and seedlings to examine the controls on germination and seedling survivorship (Angers-Blondin et al 2018). It is also easy, at the other end of the demographic spectrum, to conduct field and aerial surveys of mature or emergent shrub biomass and abundance (Tape et al 2006, Lantz et al 2013, Myers-Smith and Hik 2018) to examine correlations with disturbance and climate. Pulses of recruitment can be determined from age distributions of adult shrubs derived from dendroecological approaches (Boulanger-Lapointe et al 2014, Büntgen et al 2015, Myers-Smith and Hik 2018, Andreu-Hayles et al 2020). However, there remains a demographic gap, and difference in outcome metrics, among studies, between seedling survivorship on the one hand and growth of mature shrubs on the other. Thus, we suggest there is a need for future research on seed dispersal, and processes that control shrub success between initial establishment (i.e. <5 years) and reaching mature individuals, such as seedling competition for light and nutrients, all of which may limit long-term shrub establishment success as well as growth.
5.4. Research gap: wildfire effects on shrub expansion
While the literature clearly documents effects of wildfire on shrub biomass, litter layer, and active layer depth (see section 3.6), many gaps remain in understanding and predicting the impacts of wildfire and other disturbances on shrub expansion. Recent severe fires and repeated burns in the Arctic prompt the need to further study the effects of fire intensity and frequency on shrub expansion. For example, the effects of more severe fires on soil fungi reduced seedling performance in an Alaskan site, raising the possibility that fire–fungal–plant interactions may counteract positive aspects of fire on establishment in tundra (Hewitt et al 2016).
Ecosystem interactions can reduce or amplify the effects of fires on shrubification. For example, shrub density may be increased by wildfire and in turn shrub biomass provides fuel for fires which can promote further shrub colonization (Higuera et al 2008, Bret-Harte et al 2013). Likewise, interactions among shrub canopy, the moss layer, and permafrost thaw can dampen or exacerbate the effect of fire on active layer depth perturbations, leading to more or less shrub colonization and growth. One consequence of the importance of these internal interactions is that tundra fires may have opposite influences on shrubification depending on antecedent conditions, hydrology (e.g. drainage and slope), and nitrogen availability.
It is difficult to conduct whole-system manipulation or observational studies to determine the ultimate net effect of fire on shrubs because of the long time scales and multiple processes involved, and difficulty maintaining adequate control or untreated systems (Bouskill et al 2020). An alternative approach is to conduct more narrowly-aimed experiments to quantify the response or effect size for separate components of the ecosystem response (such as effect of fire on seed viability or seedling survival), and integrate what is learned in process-rich models. Ideally, these experimental approaches would allow for replication and evaluation in different landscapes. Moreover, model sensitivity analyses could help prioritize which processes to study.
5.5. Needed model processes, parameters, and benchmarking
In this section, we describe modeling needs most relevant to simulate changes in vegetation composition that lead to tundra shrub expansion and alter the ecosystem carbon balance. Although models vary in structure and parameterization, we highlight below the key process representations and modeling needs such as (a) tundra PFT traits, (b) topography, hydrology, and thermal dynamics, and snow–shrub interactions, and (c) synthesized observations for model benchmarking.
5.5.1. Model representation of tundra PFT traits and mechanisms
We advocate that competition among PFTs should be modeled from differences in key plant traits known to vary among species. The relevant traits that control tundra PFT competition for light, water, and nutrients are critical to accurately represent vegetation dynamics in land models. For example, light competition in many models include a light extinction coefficient and apply Beer's Law, and/or an estimate of foliar percent cover, applied to each PFT in an ecosystem (Euskirchen et al 2009, Druel et al 2019). Some Earth System Model (ESM) 'big leaf' land models (e.g. CLM (Lawrence et al 2019), ELM (Zhu et al 2019)) ignore light competition between PFTs. The light extinction coefficients are typically static in models due largely to a lack of field measurements (Zhang et al 2014). However, these coefficients may in fact change over time, particularly in expanding shrub tundra ecosystems. Consequently, for these types of models, additional data on this model parameter may be needed to more accurately simulate changing competitive interactions among PFTs for light in Arctic tundra. Other models (e.g. ecosys (e.g. Grant et al 2019a), ELM-FATES (Holm et al 2020)) explicitly represent leaf areas aggregated from those of all PFTs and resolved into multiple canopy layers. Leaf areas within each layer are then used to calculate interception of direct and diffuse irradiance by each PFT (Grant et al 2017a). In this type of model, observations of leaf reflectance, transmittance, angles, LMA, and clumping are needed for parameterization.
Traits for nutrient acquisition include, for example, maximum rooting depth, nutrient uptake kinetics that control root nutrient acquisition, and root growth that can lead to asynchrony with aboveground growth (Zhu et al 2016). This type of information is scarce and current ESM land models often do not represent the relevant wide range of PFT-specific root structural and functional properties (Warren et al 2015), although some progress is being made (Riley et al 2018, Zhu et al 2019). These distinctions may be important, for example, since more productive species, such as shrubs, use the most abundant nitrogen forms, while less productive species use the less abundant forms (McKane et al 2002). Some models may integrate these observations directly, while others may explicitly account for the underlying mechanisms and use the observations as benchmarks (Mekonnen et al 2016). Tundra field studies are improving understanding of tundra root functional traits, including, for example, luxury consumption of nutrients in fertilized tundra (van Wijk et al 2003), root nitrogen uptake under the stress of microbial competition (Zhu et al 2016), and differences in nitrogen uptake strategies between graminoids and shrubs (Wang et al 2017), thereby developing empirically based datasets of root functional traits (McCormack et al 2017) that can be used to formulate and parameterize models.
Below we highlight nine groups of PFT traits known to affect shrub growth and expansion via resource acquisition and allocation that drive growth, internal plant carbon and nutrient cycling and retention, litterfall, and light capture. We note that identifying relevant traits is important, but models must apply these traits in a robust numerical framework that allows for the effects of these traits to be expressed in plant and microbial function (Tang and Riley 2018). Our descriptions here are derived from processes included in several existing models (e.g. ecosys, ED2 (Medvigy et al 2019)).
5.5.1.1. Leaf mass: area ratio (LMA)
Leaf mass per area (LMA) is an emergent PFT property arising from changes in leaf area vs changes in leaf mass during leaf growth. To represent PFT competition for light, vertical profiles of canopy leaf area for each PFT need to be calculated, preferably from plant nonstructural C, N, and P allocation to each organ. A tractable approach to prognose direct and diffuse irradiance interception is to aggregate PFT leaf areas in each canopy layer for each model time step (Grant et al 2019b, Medvigy et al 2019). An important trait in this regard is the relationship between changes in leaf area and changes in leaf mass. For example, needleleaf PFTs have greater LMA than do broadleaf PFTs (Wright et al 2004, Serbin et al 2019). Models should represent traits that control leaf morphology and phenology to accurately simulate canopy light interception and thus PFT competition for light.
5.5.1.2. Vertical growth
PFTs differ in their allocation to vertical growth, and these distinctions are needed to represent the competitive light environment (Fisher et al 2018). For example, tall woody plants often have a larger investment in vertical growth per mass allocated than low-lying vegetation (Klein and Hoch 2015). Vertical biomass accumulation also raises maintenance respiration requirements, gradually reducing respiration available for growth (Ryan and Waring 1992). Higher vertical growth of shrubs, and longer sheaths or petioles and internodes in graminoids and forbs may enable these PFTs to gradually dominate bryophytes (Elmendorf et al 2012a). Therefore, models should include traits that control mechanisms of belowground and aboveground resource allocation and vertical elongation to accurately model PFT height and thus competition for light.
5.5.1.3. Leaf N and P contents
Needleleaf vs broadleaf shrubs have smaller leaf structural N:C and P:C ratios (Wright et al 2004, Knyazikhin et al 2013), requiring a lower parameter value for maximum leaf structural N:C and P:C ratios. This lower value is associated with the lower parameter value for leaf area growth vs leaf mass growth that causes larger LMA, as observed in the meta-analysis by Wright et al (2004). Despite larger needleleaf LMA, lower structural N:C and P:C ratios reduce their N and P densities per unit ground area relative to those of deciduous shrubs and herbaceous plants, reducing their ability to develop rapid photosynthetic rates under favorable growing conditions (Aerts 1995, Walker et al 2014). Consequently needleleaf shrubs are less competitive in warmer climates in which rapid photosynthetic rates are sustained by more rapid nutrient mineralization and uptake in warmer soils. Therefore, models should include PFT traits that control leaf nutrient concentrations, demand, and rates of uptake to accurately represent photosynthetic and respiration rates.
5.5.1.4. Leaf photosynthetic and respiration rates
Leaf carboxylation rates are driven by light and dark reactions that depend on specific activities and surficial densities of rubisco and chlorophyll (Farquhar et al 1980). Models represent these rates in different ways. In ecosys, e.g. specific activities of rubisco and chlorophyll are calculated from maximum values which can be reduced using prognosed nonstructural N:C and P:C ratios (Grant et al 2017b). Information on these maximum activities of rubisco and chlorophyll, their sensitivity to stoichiometry, and how they vary across PFTs are therefore important (e.g. Farquhar et al 1980b). Leaf maintenance respiration is affected by leaf N content, and leaf growth respiration is driven by nonstructural C, N, and P, so that PFTs with higher leaf N content have higher leaf respiration rates (Wright et al 2004, Walker et al 2014). Similarly, stem and root maintenance respiration are affected by stem and root N contents, imposing a greater maintenance respiration requirement on PFTs with large stems and roots (Stockfors and Linder 1998). Given the dynamic nature of plant organ stoichiometry, models should prognose dynamic structural and nonstructural C:N:P ratios using CO2 fixation, root N and P uptake, and allocation within the plant. In this context, traits of rubisco and chlorophyll densities and structural stoichiometry are also needed.
5.5.1.5. Leaf lifespan and turnover
Litterfall allows each PFT to carry only the leaf mass maintainable under the environmental conditions in which it is growing (Reich et al 1997). For instance, in ecosys leaf litterfall may occur whenever requirements for maintenance respiration exceed respiration of nonstructural carbon (Grant et al 2019a). In deciduous PFTs, models should also consider phenologically driven litterfall (Grant et al 2009, Grant et al 2012a, Oberbauer et al 2013), although drivers that trigger leaf emergence and leaf senescence in tundra ecosystems remain uncertain (Myers-Smith et al 2019b). Models should also represent the greater nutrient conservation of evergreen PFTs, which benefits them in nutrient limited environments such as the tundra (Aerts 1995). In general, models should represent traits that control internal carbon and nutrient cycling and retention, and litterfall.
5.5.1.6. Leaf clumping
Models should consider PFT variation in leaf clumping, since it is important in canopy radiation capture (Jiao et al 2018). Needleleaf PFTs generally have lower clumping indices (greater self-shading) than do broadleaf PFTs (He et al 2016). These indices determine the fraction of leaf area index exposed to direct and diffuse irradiance by each PFT in each canopy layer and hence relative dominance of taller over shorter canopies.
5.5.1.7. Root growth
Models should represent PFT-specific dynamic root length and density since these traits affect nutrient and water uptake (Grant 1993, Zhu et al 2016). PFTs with large populations and hence small individual size (e.g. bryophytes) have smaller primary root extension and hence more shallow root systems, while PFTs with small populations and hence larger individual size (e.g. shrubs) have greater primary root extension and hence deeper root systems, although the depth may be constrained by permafrost. Deeper root profiles improve competitive access to deeper water and nutrients, but may reduce root density at shallower depths, reducing competitive access to shallower water and nutrients (Schimel et al 1996, Wang et al 2018, Hewitt et al 2019). These differences in root profiles affect competition for soil water and nutrients among shallow- vs deep-rooted PFTs. Models should represent traits that control root morphology, phenology, and growth to accurately predict root length, radius, and density and thus nutrient and water uptakes.
5.5.1.8. Root hydraulic resistance
Root axial resistivity governs water uptake along soil–root–canopy hydraulic gradients (Welegedara et al 2020). Needleleaf PFTs have higher root axial resistivity than deciduous PFTs (Larcher 2003, Maherali et al 2004, Anderegg 2015). Higher root axial resistivity results in slower water uptake and hence lower canopy water potential and stomatal conductance, while lower stem and root axial resistivities improve water uptake, and thus increase water potential and stomatal conductance. Slower water uptake conserves water, providing protection from cavitation, making ecosystems dominated by needleleaf PFTs less sensitive to short-term droughts (Wang et al 2002, Grant et al 2009), but may disadvantage needleleaf PFTs in warmer climates with greater transpiration demands and hence more rapid water uptake needs. Increased stem resistance forces lower canopy water potentials and hence stomatal conductance, reducing CO2 fixation and hence dominance (Grant et al 2017). Thus, traits that affect root and stem hydraulic resistances need to be considered in models to simulate differences in rates of water uptakes among PFTs.
5.5.1.9. Root porosity
PFTs adapted to saturated soil moisture conditions, such as sedges, have larger aerenchyma and hence can transfer O2 more rapidly to root respiration sites when gaseous O2 diffusion through soil is limited by low air-filled porosity (Grant 1993, Pan et al 2019, 2020). Therefore, larger aerenchyma fractions allow for increased root respiration, growth, and nutrient and water uptake under saturated conditions (Grant et al 2012b, Pan et al 2019, 2020). Smaller root porosity fractions of shrubs therefore reduce their competitive ability in wetlands. Thus, models need to represent PFT-specific root porosity traits.
5.5.2. Research gap: model representation of tundra PFTs
To capture how shrub expansion may influence other tundra PFTs and ecosystem function, models of shrub expansion need to consider shrubs and other vascular and non-vascular vegetation. Early PFT classifications, such as those proposed by Chapin et al (1996a), recommend the use of several tundra-specific PFTs, including deciduous shrubs, evergreen shrubs, sedges, grasses, forbs, Sphagnum moss, non-Sphagnum moss, and lichens. Models that have included PFT groupings similar to those recommended by Chapin et al (1996a) have simulated how projected shrub expansion may influence other PFTs. For example, shrub expansion reduced moss growth due to shading effects (Epstein et al 2000, 2001, Mekonnen et al 2018b), which then has implications for changes in the soil thermal regime and active layer thickness (Lawrence and Swenson 2011). However, these PFT groupings were shown to explain variation in traits related to resource-economic but not size-related traits (Wullschleger et al 2014, Fisher et al 2018), suggesting the need for a new classification and representation of tundra plants in land models.
Models may also need to consider several types of deciduous shrubs, including tall shrubs such as alder (e.g. A. viridis spp.), birch (Betula spp.) and willow (e.g. Salix spp.). For instance, tall willow and alder shrubs, typically found along hillslopes and stream beds, may trap more snow, resulting in the development of taliks (perennially thawed ground within permafrost; Pomeroy et al 2006). Tall alder shrubs may also decrease diversity of other tundra plant species through modification of the local environment by capturing snow, high rates of transpiration, and nitrogen fixation (Wallace and Baltzer 2020). The inclusion of birch vs willow shrubs is important since it has implications for animal habitat of some large herbivores. For example, in tundra, moose preferentially browse willow shrubs, and in particular, in winter, browse willow shrubs taller than 1 m since these taller willows typically remain exposed above the snow (Tape et al 2016). Other animals (e.g. reindeer) also feed on non-vascular vegetation, affecting competitions with shrubs.
5.5.3. Model representation of tundra topography, hydrology, and thermal dynamics, and snow–shrub interactions.
Most land models (including those in ESMs) do not account for the underlying mechanisms for three-dimensional landscape hydrological dynamics; fine-spatial scale topographically driven surface and sub-surface nutrients and energy flows; thermokarst and subsidence (Nelson et al 2001, Turetsky et al 2019); and the effects of microtopographic and sub-gridcell scale soil heterogeneity (Grant et al 2017c, Bisht et al 2018). As a result, widely observed shrub growth and expansion hotspots (e.g. hillslopes and terraces (Naito and Cairns 2011, Ropars and Boudreau 2012)) may not be accurately represented. Therefore, land models need to account for coupling of surface and sub-surface lateral interconnectivity in topographically diverse tundra landscapes to predict current and future distributions of shrubs across the Arctic.
To accurately simulate seasonal nutrient and energy cycles, land models should also account for the effects of topography and wind on snowpack redistributions across a landscape (Essery and Pomeroy 2004). Multi-scale field natural and manipulation (e.g. snow-fence) experiments (Wipf and Rixen 2010) are required to better understand, inform, and test models regarding the impacts of shrubs on snow accumulation, albedo, timing of snowmelt, phenology, soil temperature, and litter decomposition rates. Models should also represent processes following wildfire such as effects on permafrost thaw and thermokarst development (Schuur et al 2007, Jones et al 2015) and its subsequent effects on soil moisture and thermal regimes, nutrient cycling, recovery patterns, and carbon cycling.
5.5.4. Development of synthesized observations for model benchmarking
The wide range of processes affecting shrub dynamics (figure 1) imply that multiple combinations of processes could lead to comparable (i.e. within observational error) emergent shrub biomass, complicating model evaluation. Therefore, observations for model testing are needed to test individual processes in addition to emergent responses. We suggest the mechanisms discussed in section 3 be used to develop individual-process and emergent benchmarks. The role of nutrient dynamics or warming can be quantified using fertilization experiments (e.g. Shaver and Chapin 1980, Mack et al 2004, Bouskill et al 2014). Plant–plant competition benchmarks can be developed from exclusion experiments (e.g. Hobbie et al 1999, Rixen and Mulder 2009). Hydrological benchmarks can be developed from rain exclusion or watering experiments (e.g. Keuper et al 2012). Disturbance benchmarks can be developed from intentional manipulations (e.g. Schadel et al 2018) or natural disturbances (e.g. Mack et al 2011, Iwahana et al 2016). Further, since short- and long-term ecosystem responses can differ substantially (Bouskill et al 2020), we advocate for model benchmarks that explicitly account for time. Finally, valuable benchmarks include those that consider perturbation responses, either to experimental manipulation or other forcing changes (e.g. precipitation, fire). Benchmarking can augment the traditional approach of evaluating a model against observations of an emergent response that depends on many competing, and difficult to measure, factors (Collier et al 2018).
6. Conclusions
Rapid climate warming is causing substantial changes in ecosystem structure and function of northern ecosystems. Results from in-situ observations, warming experiments, and remote-sensing vegetation indices have shown that woody shrubs are expanding across the circumpolar Arctic (Cornelissen et al 2001, Tape et al 2006, Myers-Smith et al 2011, Elmendorf et al 2012b, García Criado et al 2020). Modeling studies also show that this recent shrub expansion will continue under the projected warmer Arctic (Euskirchen et al 2009, Mekonnen et al 2018b). A shift in vegetation composition that results in a woodier Arctic will alter the ecosystem carbon balance by affecting a complex set of soil–plant–atmosphere interactions (Elmendorf et al 2012a, Loranty and Goetz 2012, Mack et al 2004, Weintraub and Schimel 2005, Parker et al 2021). Much of the tundra is underlain by permafrost that stores large amounts of frozen carbon (Brown and Romanovsky 2008, IPCC 2013, Hugelius et al 2020, Turetsky et al 2020). Shrub expansion can alter the seasonal surface energy balances leading to deeper active layers and thus expose this permafrost carbon to microbial decomposition (Blok et al 2010, Lawrence and Swenson 2011, Nauta et al 2015). These processes have a wide range of implications for net ecosystem carbon exchange and associated feedback to the climate system (Blok et al 2010, Bonfils et al 2012, Loranty and Goetz 2012, Lafleur and Humphreys 2018).
Our review indicates that the dominant controlling mechanisms for shrub biomass increases include: (a) climate warming, (b) soil moisture and snow dynamics, (c) topography, (d) permafrost thaw, (e) nutrient dynamics, (f) disturbance (e.g. wildfire), and (g) herbivory and grazing, and interactions among these factors. We showed that the mechanisms through which Arctic shrub expansion alters ecosystem carbon balance are complex and involve interactions among several biotic and abiotic factors (figure 1) such as warming, surface energy balances, snow–shrub interactions, permafrost degradation, litter inputs, and nutrient cycling (Cornelissen et al 2001, Sturm et al 2005a, Myers-Smith et al 2011a, Elmendorf et al 2012a, DeMarco et al 2014a, Martin et al 2017). Our review motivated our recommendations for model mechanisms required to accurately predict future shrub expansion rates. Although there have been a number of field experiments to characterize the impacts of climatic drivers (e.g. air- and soil-warming experiments) on shrub expansion, warming often covaries with soil moisture, snow dynamics, active layer depth, nutrient availability, and other environmental controls that directly or indirectly affect shrub growth and establishment (Elmendorf et al 2012a, Martin et al 2017). Thus, multifactorial field experiments and multi-scale observations, designed to disentangle the relative impacts of multiple drivers and their interactions, are needed to better understand the mechanisms of tundra shrub expansion under changing climate. Model process representations should be informed by and tested against field experiments and observational studies spanning multi-spatiotemporal scales.
Acknowledgments
This research was supported by the Director, Office of Science, Office of Biological and Environmental Research of the U S Department of Energy under contract DE-AC02-05CH11231 to Lawrence Berkeley National Laboratory as part of the Next-Generation Ecosystem Experiments in the Arctic (NGEE-Arctic) project (Z.A.M, W.J.R, N.J.B, M.S.T, and Y.L), NASA ABoVE (Arctic Boreal and Vulnerability Experiment (Grant No. NNX17AC57A to G.I and 80NSSC19M0112 to S.J.G and L.T.B) and NERC (Grant No. NE/M016323/1 to I.M.-S).
Data availability statement
All data that support the findings of this study are included within the article or references provided in the figure captions (and any supplementary information files).