
Abstract
Nature is full of exemplary species that have evolved personalized sensors and actuating systems that interface with and adapt to the world around them. Among them, cephalopods are unique. They employ fast-sensing systems that trigger structural changes to impart color changes through biochemical and optoelectronic controls. These changes occur using specialized optical organs that receive and respond to signals (light, temperature, fragrances, sound, and textures) in their environments. We describe features that enable these functions, highlight engineered systems that mimic them, and discuss strategies to consider for future cephalopod-inspired sensor technologies.
Export citation and abstract BibTeX RIS

This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 License (CC BY, http://creativecommons.org/licenses/by/4.0/), which permits unrestricted reuse of the work in any medium, provided the original work is properly cited.
Many biological systems have evolved advanced sensing features to interface with the world around them. These systems may receive, respond, and distribute optical, chemical, physical, or electrochemical signals to impart a physical or chemical change, typically in a closed loop. 1–4 Regardless of their specific operational mode, a common thread of natural sensors is their utility of the G-protein coupled receptor (GPCR) superfamily of proteins. These transmembrane proteins consist of seven α-helices connected by complementary loop regions and exist in a dynamic equilibrium between inactive and active states. The binding of specific agonists to the extracellular portion of this biomolecule shifts equilibria towards the activated states, which cause conformational changes and elicit additional signaling events. 5 These precipitated signals ultimately lead to the activation of other proteins and/or the production of secondary signaling molecules that impart a corresponding optical, chemical, physical, or electrochemical response. 6,7 With the exception of gustatory detection of sour and salty taste sensations 6 and mechanical sensors used in tactile perception, 8 which both use specialized ion channels to propagate action potentials, GPCRs are used in most natural sensing and actuation events in biology.
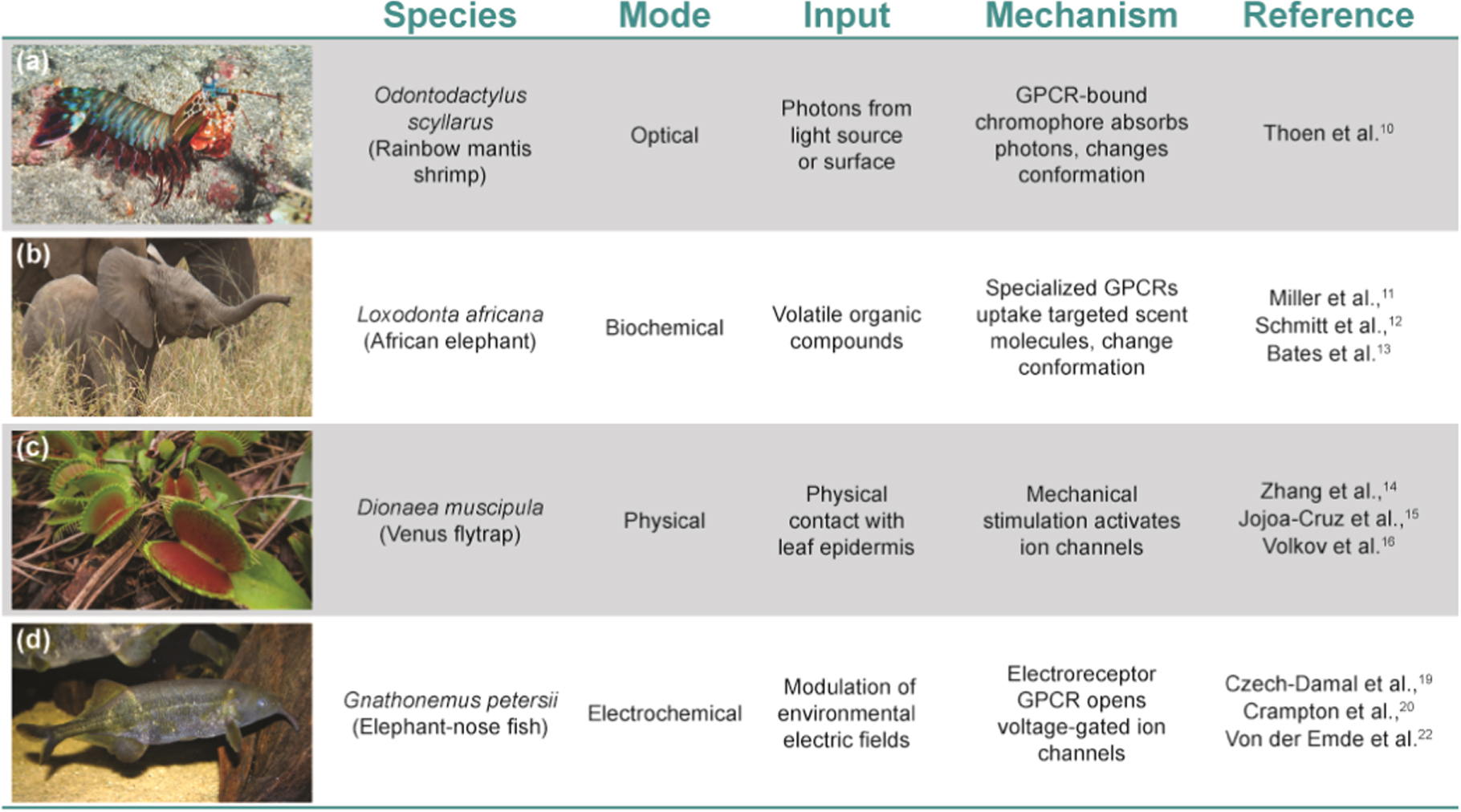
Natural sensors/actuators are usually localized to specific cells of organs in living systems. Ocular photoreceptive organs are an example of sensors that are present in almost all living systems but vary dramatically across species, ranging from the highly tunable single-unit eyes of mammals to the collected ommatidia common in the insect world. 9 The mantis shrimp represents one extreme example with the most diverse array of known photoreceptors that can recognize (although not distinguish between) colors (Fig. 1a). 10 Other examples include olfactory sense organs which detect volatile organic compounds (VOCs) and have been optimized for certain species like the African Elephant with >2000 receptor genes 11 used in feeding, 12 evasion, 11 and herding (Fig. 1b). 13 Mechanical sensors like those found in the sensory hairs of the Venus flytrap (Dionaea muscipula) are another example used to sense prey via mechanosensitive ion channels that can then trigger turgor pressure and hydrodynamic flow to close their trap within ∼100 ms (Fig. 1c). 14–18 Finally, natural electrochemical sensors like those found in the elephant-nose fish species Gnathonemus petersii incorporate specialized receptor cells called electrocytes which detect differential electric field gradients and activate voltage-gated sodium channels to transduce signals through to the central nervous system (Fig. 1d). 19–21 In all cases, the sensitivity of the specific receptors varies across species, depending on the density and arrangement of the receptor cells in their respective organ systems. In some cases, signals generated by the animal's own electrolytes can also be adapted for communication and navigation. 21,22

Figure 1. Categorization of living biological sensors that utilize specialized GPCRs. a) The mantis shrimp is equipped with complex eyes with 12 different photoreceptor types. b) African elephants use their trunk to smell their surroundings. c) The Venus fly trap utilizes its open lobes to close abruptly with a mechanical trigger. d) The elephant-nose fish is capable of electrically imaging the external environment around them. O. scyllarus image copyright Jean-Paul Cassez 75 ; L. africana image copyright Jean-Paul Boerekamps 76 ; D. muscipula image copyright Miguel Vieria 77 ; G. petersii image copyright Brian Gratwicke. 78 All images adapted under Creative Commons license (CC BY-NC).
Download figure:
Standard image High-resolution imageThese examples represent only a snapshot of biological systems that utilize an adaptive biosensing scheme to perform a specialized optical, chemical, physical, or electrochemical task. In these cases, however, such schemes generally incorporate only one of these modes for specialized sensing. Unlike the other systems discussed above, cephalopods incorporate multi-modal sensing and signaling sometimes simultaneously to interface with the aquatic world around them (Fig. 2). 14,23–26 While the exact mechanisms regulating these sensing interactions are still unknown, data suggest that cephalopods exist as living analytical sensors with discretized sub-sensor components capable of computing complex tasks to display patterns of incredible resolution. In this perspective, we describe the fundamental components regulating the complex cephalopod sensory system, introduce key technologies inspired by them, and highlight the opportunity for further exploration/inspiration to be considered in the design of future materials.

Figure 2. An Octopus vulgaris receives various stimuli to inform its creation of an adaptive display. (a) Stimuli include (i) light from the environment as detected through the use of ocular and extra-ocular light receptors, (ii) water-soluble environmental compounds that passively diffuse into the environment from predator and prey species as well as potential mates that are detected through several chemoreceptive organs, olfactory pits, and gustatory tissues, and (iii) physical stimuli like temperature and contact-pressure, as well pseudo-auditory vibrations which are detected by sense organs called statocysts. (b) Incoming signals are processed in the nervous system, which includes the brain and associated neural networks. (c) Adaptive display is enabled by a multifarious and stratified organization of tissues. E, epidermis; DM, dermal erector muscle; YC, yellow chromatophore; RC, red chromatophore; BC, brown chromatophore; I, iridophore; L, leucophore. Octopus vulgaris image copyright Callum Evans, adapted under a Creative Commons license (CC BY-NC). 79
Download figure:
Standard image High-resolution imageCurrent Status
Cephalopods have a remarkable visual sensing system that includes their ability to adjust ocular focus rapidly, allowing them to see objects clearly at multiple distances. 27–29 Cephalopod eyes also have a high density (90,000 cells mm−2) 30 of photoreceptor cells, allowing them to perceive fine details and colors, as well as the ability to register polarized light due to specialized retinal cells. These features help the animals to navigate in the ocean and locate prey. Additionally, cephalopods have a remarkable ability to sense their environment through chemoreception, which enables the animal to detect water-soluble biochemicals in a process analogous to smell or taste. These chemoreceptors, which in the case of octopuses are located on their suckers, the buccal lips surrounding their beaks, and in their olfactory receptor organs, allow the animal to detect a wide range of chemical signals, identify the presence of prey, and avoid predators. 31 Cephalopods can also use these biochemical detection methods to communicate with one another. For example, octopuses release a chemical signal when they are threatened or attacked, which can attract other octopuses to help defend them. 1,32 Cephalopods are also equipped with specialized sensory structures called statocysts that allow them to detect pressure changes in the water. Statocysts are fluid-filled sacs that contain a small, dense object called a statolith. 33 When the cephalopod moves or the water pressure changes, the statolith moves within the fluid in the statocyst, stimulating sensory cells that then transmit information to the cephalopod's brain to determine the direction and magnitude of the pressure changes. In addition to detecting pressure changes, statocysts also play a role in the cephalopod's sense of balance and orientation in the water, allowing the animal to navigate its environment and avoid predators. 34,35
Another notable feature of cephalopods is their advanced display ability that allows them to survive in their environment and interact with their surroundings in remarkable ways. For instance, they can adapt their skin color and, in some species, texture based on their mood or environment, allowing them to blend in and avoid predators or surprise prey. 31,36 Their complex brain, specifically their highly developed optic lobe located behind their eyeballs, 37 regulates this process. This part of the brain receives and processes visual, biochemical, and physical information to aid in the perception, selection, and adaptive control of color and pattern changes in their own skin. It has been reported that cephalopods can generate over 5000 different skin patterns using a combination of their dermal optical organs. 25,38 Although the complete mechanism underlying pattern generation is still unknown, it is understood that brain cells in the optic lobe select an appropriate body pattern from the set of patterns registered in the central nervous system and that specific motor neurons associated with dynamic pigmentary organs effect the visual changes. 39
These dynamic pigmentary organs in cephalopod skin are complex. They are made up of several layers of optical tissue which all contribute to the generation of a final adaptive display. 40–42 The top layer of the skin comprises a thin epidermal layer, and in benthic coleoid cephalopods such as octopus and cuttlefish, a number of dermal papillae which control the outward texture of the animal. 43 The morphology and prevalence of these papillae varies significantly between the cephalopod species, but the underlying strategy uses a combination of dermal erector muscles, which create convex outcroppings, and retractor muscles, which serve to contract these extensions. 43 Underneath this complex outer layer lies dermal tissue specifically optimized for manipulating incident light. Vertically stacked yellow, red, and brown optical filters (chromatophores) are the uppermost strata. These chromatophores, which are attached radially to dozens of muscle cells and can be expanded and contracted through innervation by the nervous system, 44,45 are activated as fast as five times per second. 46 These organs are positioned over a layer containing Bragg reflectors (iridophores) and, in cuttlefish and octopus, white light diffusers (leucophores) which are interspersed as the bottom most optical layer. 40,44,47 All of these elements can be activated to generate broad-area visible patterns to manipulate the visible color and pattern of the animal.
There are three main features of cephalopod camouflage that differentiate them from other systems in biology. The first is the degree of background matching, which includes overall pattern, intensity, and color. 36 The second is the speed of color change, which has been reported to be as fast as 100 msec. The third is 3D textural changes which are enabled in some species due to adaptive sensory papillae. 38 Using these features as a general framework, we propose four categories of cephalopod-inspired technologies (Fig. 3). The first is based on manipulating the physical form (i.e., contraction and expansion) of an active visual component to change the holistic visual appearance of a material (Fig. 3a). This method utilizes force to modify shape/size, resulting in changes in width or length either along two axes as in concentric models 48 or along one axis as in uniaxial extension. 49,50 Another approach is based on exposing color/pattern through layered structures comprising a semiflexible or fracturable optically inactive top layer that covers a second active layer (Fig. 3b). As the top layer is stretched, it unveils the active layer underneath. The second layer may consist of fluorescent/iridescent components or a concealed pattern. 51,52 Alternatively, an electrical or magnetic field can be used to rotate or align particles embedded within a material, thereby revealing a pattern or transforming a colored/opaque material into a transparent one, or vice versa. 53 A third approach to color change can be achieved through chromophore manipulation, such as through redox chemistry 54,55 or mechanochromism (Fig. 3c). 56,57 Finally, color in a material can be manipulated through adaptive structural coloration (Fig. 3d). In this method, the material is infused with crystalline substances that possess reflective properties. 56–58 By adjusting the spacing between crystals and crystalline layers, it becomes possible to modify the wavelength of incident light and generate different colors reminiscent of adaptive iridophores.

{kind=link}
{kind=link}
Figure 3. General classification of materials strategies and the biological phenomena that inspire them. Cephalopod-inspired strategies incorporate at least one of the following: a) radial or uniaxial expansion and contraction of an optically active material to recapitulate the actuation of the chromatophore, or when employed in a three-dimensional matrix, can mimic cephalopod papillae; b) adaptive visibility via the lateral expansion of layered materials revealed through the cracks; c) stimuli responsive small molecules that change color through redox activity, expansion of π-conjugation, or other means; d) dynamic structural coloration inspired by cephalopod iridophores.
Download figure:
Standard image High-resolution image{kind=link}
There have been several attempts by materials scientists and engineers to emulate one or all three of these features to create stimuli-responsive color-changing systems. 1,59–61 Inspired by the sensing and actuating iridocytes, Wang et al. fabricated a chromotropic ionic (CI) skin with the ionic and optical dual signal output to respond to multiple stimuli, including pressure, tension, infra-red (IR) light, and temperature. 58 In these systems, color-change occurs via ionic conductive hydrogels based on N,N-dimethyl-1-dodecylamine N oxide protonated surfactant molecules embedded into a cross-linked hydrogel matrix (poly-(diacetone acrylamide-co-acrylamide)). This combination gives rise to variably reflected colors when the hydrogel is stretched, effectively increasing the lattice spacing to mimic the reflective plates in iridophores. This biomimetic "skin" can produce reversible resistance signal variation and continuous color switching from red to blue under tensile strain and thermal stimuli. 58
Separately, Wang et al. designed an artificial chromatophore-inspired electro-mechano-chemically responsive (EMCR) elastomer system that can exhibit a wide variety of fluorescent patterns under the control of electric fields. 48 The EMCR elastomer was prepared using spiropyran, a mechanophore that emits fluorescent signals under mechanical stresses, embedded in a cross-linked silicone elastomer. The application of high electric field (31.6 kV mm−1) is enough to destabilize the EMCR and create a pattern of wrinkles. As the applied electric field continues to increase (63.1 kV mm−1), the patterned wrinkles become circular craters, highlighting a stimuli-responsive mode to generate color and textural changes. 48
Other approaches to replicate adaptive coloration in cephalopods include variations of light, 62,63 thermal, 49,50,64 mechanical, 51,52,65 chemical, 66–68 electrochemical, 54,55,69 or magnetic 53 control strategies in response to passive and/or active stimuli. The creation of more complex, multi-material design schemes for dual mode (both structural and pigmentary) color changes have also been explored. Clough et al. have created a device using a spiropyran-containing top layer prepared with a photonic bottom layer arrayed with silica nanoparticles. 57 Under deformation, the spacing among the periodically arrayed silica particles grows, producing a colorimetric response that can span the visible spectrum. Hsieh et al. separately created optically tunable photonic crystals inspired by cephalopods made from ordered hollow nanospheres, whose spacing varied using activated poly(3,4-ethylenedioxythiophene)-polystyrene sulfonate (PEDOT:PSS) structures to create iridophore-like Bragg reflecting structures. The resultant photonic crystal could be activated by controlling the applied voltage, causing structural colors to change between green, yellow, orange, and red at 0, −0.3, −0.7, and −1.0 V, respectively, over 100 reversible cycles. 56
Inspired by cephalopod dual function of simultaneous pressure detection and visualization, Guo et al. have fabricated a hybrid device that responds to pressure changes and generates a capacitance signal from a real-time visual color change. 70 The device is created using magnetron-sputtered tungsten oxide (WO3) on 3-D printed structures layered on an ionic gel (polyvinyl alcohol/H3PO4), which is sandwiched between two indium tin oxide (ITO) electrodes. When pressure is applied, the color of the device changes from yellow to blue due to the redox activity of W6+ to W5+/W4+. Applications of this system have extended to the development of a smart eyeglass system that is capable of monitoring physiological parameters of the human body (ex: pulse rate) and connecting to a tablet wirelessly to monitor changes in real-time, demonstrating the power of alternative applications of cephalopod-inspired technologies. 70
Conclusions and Future Perspectives
While most cephalopod-inspired systems historically focus on replicating the fast and adaptive color-changing features of the animals using external stimuli like temperature, pressure, voltage, or strain, there are several unexplored applications that warrant materials development. For example, cephalopods have complex eyes with excellent vision, 27,28 as well as sensitive hair cells that detect vibrations and changes in water pressure. 33–35 Understanding the structure/function of these systems could provide a fascinating source of inspiration for the development of new types of sensors that could have a wide range of applications, from environmental monitoring to medical diagnostics.
While the potential for such cephalopod-inspired technologies is extraordinary, the reality is that the system is overwhelmingly complicated by the intricate innervation, complex coupling of cells across multiple tissues and organs, and the inherently adaptive feature of cephalopod skin. Advancements in the field of genetics have allowed scientists in the past decade to begin uncovering the gene sequences of cephalopod species, such as the California two-spot octopus (Octopus bimaculoides), 71 the Hawaiian bobtail squid (Euprymna scolopes), 72 and the Pharaoh cuttlefish (Sepia pharaonis) 73 to begin tracing the molecular origins of their functions. As recently as 2020, the first transgenic cephalopod model was developed by Crawford et al. in the squid Doryteuthis pealeii, allowing for knockout and functional investigation of specific gene sequences. 74 As these genetic data are compiled, a concerted effort must be made to interpret and annotate them to draw meaningful conclusions of their functional roles in the animals. Initial attempts to contribute to this effort have already started in recent investigation of the structural and compositional analysis of optical organs such as the chromatophore to better understand how these systems function 42 but more work is needed to connect all the component parts together to better inform new applications.
The current limitations of the materials development space largely stem from the natural focus on display end-applications. While these technologies offer significant advancements for new optical materials, little work has been done on advancing adaptive and/or regenerating sensors that may drive communicative color components based on the cephalopod's natural operational modes (optical, biochemical, and physical). Most current contributions are instead based mostly on mechanical actuation, thermal, and/or electrical activation using commercial off-the-shelf components. To bridge this gap, the development of autonomous distributed sensors that sense color/patterns in the environment then deliver the information throughout adaptive systems should be developed. Given the complexity of this task, it will require a concerted effort among materials scientists, chemists, engineers, and biologists to make meaningful advancements in connecting the compositional contributions to structural and functional roles describing cephalopod camouflage. This advancement requires a more nuanced understanding of the component parts underlying cephalopod mechanics, demonstrating a clear need for dedicated study into the fundamental science of cephalopod biology, particularly those elements tied directly to their adaptive sensing-display feedback loop. This fundamental understanding will not only pave the way for other researchers to merge these approaches but also facilitate the creation of advanced materials for the future of cephalopod-inspired technologies.
Acknowledgments
This work was supported in part by the Office of Naval Research (Award N00014–22–1–2053) and National Science Foundation (Award DMR-1712345).