
Abstract
Coordination comes in many guises and represents one of the most striking, but least understood features of living things. The different forms that coordination takes and how it emerges and changes are of great interest to many disciplines, particularly the social and behavioral sciences, neuroscience, psychology, biology and physics itself. Inspired originally by theories of self-organization in open, nonequilibrium systems, the science of coordination (coordination dynamics) seeks to understand coordinated patterns at many scales and for multiple functions in living things. Here we review some of the key concepts of coordination dynamics along with some recent developments and posit ten tenets of the theory that may guide further understanding.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
'The power of life and will by which animals move their bodies with great and lasting force... demonstrate that there has to be other (undiscovered) laws of motion'
Isaac Newton, Letter to Oldenberg, 1675
'Physical science is that department of knowledge which relates to the order of nature... The name is applied to those branches of science in which the phenomena considered are of the simplest and most abstract kind, excluding the consideration of the more complex kind, such as those observed in living things'.
James Clerk Maxwell, Matter and Motion, 1877
"What is this 'I'?"
Erwin Schrödinger, What is life? 1944
Introduction
This paper is the third in a trilogy of articles on the science of coordination, coordination dynamics. The forerunners of the present piece dealt with connecting empirical findings and theoretical models of coordination in small and large groups of agents (Kelso 2021a) and addressed the broader impact of the elementary Haken–Kelso–Bunz (HKB) model paradigm for the behavioral, brain, developmental and social sciences (Kelso 2021b). The kinds of phenomena addressed here deal largely with basic forms of human sensorimotor control and coordination, particularly, though by no means restricted to, rhythmic kinds of behavior that are observed at multiple levels of description, from the cellular to the social.
The aim of the present perspective paper is to present some of the main concepts, methods, and tools of coordination dynamics for those in the interdisciplinary field of complex systems interested in understanding the coordination of living things. Following the quotes above from Newton, Maxwell and Schrödinger, the perspective taken here may be considered quite radical, if not heretical. It follows a line of argument by the late Robert Rosen (1991) which says that contemporary physics is too special to accommodate the class of material systems called organisms. Put another way, according to Rosen, the material systems that express the functional order of living things are as, if not more general than those that physics currently addresses. Even if one is skeptical about this, at the very least Rosen points to an extension of physics as we know it. Rosen's detailed arguments boil down to the fact that biological systems are too complex to be fractionated. This is hardly news to the complexity scientist, whose familiar mantrum is that the whole is greater than (and different from) the sum of its parts. But how can this be? The fact that we use the word 'parts' in the first place (and synonyms such as elements, constituents, components, etc) implies that nature may be broken down into parts that somehow, at least in the case of living things, are brought together to produce organized wholes. A challenge for complex systems then, is said to lie not only in elucidating the parts and their connections but also how such connections constitute an entirety. Rosen's viewpoint, however, is even more radical: he argues that mechanism and the machine metaphor do not work if the goal is a true science of complexity and must be discarded. The reason he gives is that not only are organisms complex; their organization inherently involves functions and the interrelations of such. There is no one to one relationship between such relational organizations and the structures that realize them. Such basic differences between organisms and machines (and their syntactic models), according to Rosen, force a reconsideration of what kind of science is needed to understand life.
With Newton's remarkable 'confession', Maxwell's separation of the inanimate (simple) and the animate (complex), Schrödinger's provocative question and Rosen's admonitions about 'life itself' in mind, the present paper addresses several aspects of the science of coordination. With a primary focus on the behavior of human beings, in the following we consider a number of conceptual and empirical developments of coordination dynamics (including one of its technological consequences):
- (a)The foundational notion of synergy, hypothesized to be the basic functional structure of living things and proposed as nature's way to handle complexity;
- (b)Our current theoretical understanding of how synergies are formed, how they transform smoothly and change spontaneously to accommodate current conditions, how synergies can co-exist, how new synergies can be created as others disappear, and so forth. Some of the key concepts of synergetics and the tools of nonlinear dynamical systems will be invoked, such as order parameters or collective variables, symmetry breaking, dynamical stability and instability, familiar to physicists but now shown—by means of mappings between mathematical models and empirical data—to be central for understanding biological coordination;
- (c)How the foregoing concepts and phenomena led to the development of ideas central to coordination dynamics such as intrinsic dynamics and functional information;
- (d)The discovery and centrality of (symmetry breaking) metastability, the complementary tendency of complex systems such as the brain to display both interdependence among its elements (integration) and a capacity for those elements to express themselves independently (segregation);
- (e)Supporting its utility, the realization of coordination dynamics in an application—a principled approach to human ∼ machine interaction called the human dynamic clamp—which is proving useful to understanding real-time social coordination; and
- (f)In response to Schrödinger's question, how recent work on human infants embedded in a specific environment is revealing how spontaneous self-organizing processes give rise to the 'self' or the directedness property of complex, biological systems, usually contained in terms like 'agency' 'purpose' and 'intention'. We conclude with a summary of the main theoretical tenets of coordination dynamics, a theory of directed self-organization, intended to provide a stimulus for further research.
1. Why synergy?
'Synergies... in which very few controls can manipulate a much larger number of configurational degrees of freedom... are everywhere in biology, as they are in any constrained mechanism. In biology they are indicators of complexity rather than mechanisms under constraints' (Rosen 1991, p 271).
In general, synergy refers to the combined effects that arise from interdependence among parts and processes in a given context that are not possible or achievable from those entities acting alone. A strong case can and has been made for functional synergies as the drivers for the evolution of cooperation in complex systems. Peter Corning (1983) for example has referred to this as the 'synergism hypothesis', and the biologist John Maynard Smith (1983) as 'synergistic selection' (for many examples, see Corning 2018). In the field of movement, the synergy is a functional entity: it refers to a collection of relatively independent degrees of freedom that are temporarily constrained to act as a single functional unit. This means that the very many degrees of degrees of freedom involved organize themselves in such a way as to accomplish a given task or function (see Gel'fand et al 1971, Latash 2008, Sheets-Johnstone 1999, Tuller et al 1982. For an early example in voluntary limb movements, see Kelso et al 1979). Synergies solve four main problems characteristic of complex biological systems. One is degeneracy or functional equivalence: the fact that the same outcome can be achieved using coordinated combinations of very different components. Two is multifunctionality: the fact that different outcomes can be achieved using the same components. Degeneracy and multifunctionality are ubiquitous in living things; they are observed at multiple levels of description in contexts that range from the genetic to the social (see Greenspan 2012 and Kelso 2012 for reviews). Three is how, in typical task situations, the relations among interacting components (such as their relative timing) may be preserved despite changes in parameters. Synergies have long been proposed to guarantee this 'invariance under transformation' property (Kelso and Tuller 1984 for review). Four, Rosen's quote and a focus here, the central problem of control and coordination in complex biological systems: how to compress a state space of very many dimensions into a control space of just a few dimensions. All four problems are solved by the synergy hypothesis. This also implies that the collective or group is the relevant level of description for synergistic action, a point that we will return to shortly. Synergies are what complexity scientists might call 'self-organized' which means that fundamentally synergies are the biological equivalent of a cooperative phenomenon. But is the synergy, in fact, an example of a physical cooperativity? How would we know?
Two main lines of evidence support the synergy hypothesis. The hallmark of a synergy is that any unexpected perturbation applied to the system is immediately compensated for by remotely linked members of the putative synergy in such a way as to preserve the system's functional integrity. Experiments on human speech production have shown exactly that: using a combination of novel perturbation techniques, fine wire intramuscular electrodes and continuous movement recordings, the same perturbation applied to the jaw as it lowers and raises was rapidly and remotely compensated for by different muscles in the tongue, lips and larynx depending on the specific sound the person intended to produce. That a challenge to one member of a group of potentially independent components (here the speech articulators and their neuromuscular control elements) was met—on the very first perturbation experience—by remotely linked members of the group in such a way as to preserve the speaker's intent, provides direct evidence for functional synergies or coordinative structures (Kelso et al 1984, Shaiman 1989, Sorensen et al 2019; see also Guenther et al 2006 for a specific model).
Complex systems are all about understanding indirect effects of causes. Pushing in a complex system in one place has effects in other places at some remove from the site of perturbation. The poetic image of Francis Thompson (1859–1907) comes to mind: all things by immortal power near or far, to each other linked are, that thou canst not stir a flower without troubling a star. The synergy is the epitome of a complex system. And a complex system is a synergy of synergies. When a piece of rat cortex consisting of 446 axons, 449 synapses and a small part of a single astrocyte is viewed at a nanometer scale, the complicated 3D tangle of neural and glial processes is reported to be a 'tangled mass whose topology resembles soap bubbles' (Sejnowski 2016). A synergy is like a lightly touched bubble: it ripples everywhere and trembles until its balance is restored. Though possibly underappreciated in neuroscience, this kind of 'indirect cooperativity'—an essentially non-local characteristic of synergy—has long been recognized under the term 'allostery' in studies of metabolic pathways (Monod and Jacob 1961).
A second line of evidence comes from very basic studies of sensorimotor coordination initially performed in the late 70s and early 80s in which rhythmic interlimb movements proved to be a paradigmatic window into synergy formation and change (Kelso 1981, 1984, Haken et al 1985; see Kelso 2009/2013, Turvey 2007, Newell and Liu 2020 for reviews1 ). The reason is that all the features of nonequilibrium phase transitions—considered the core of self-organizing processes in nature (Haken 1977, Nicolis and Prigogine 1989)—have been observed and modeled at both behavioral and brain levels (Kelso 2021b for recent review). As prospective 'atoms of function', synergies have been demonstrated to extend beyond the individual person (and the individual brain) to dyadic and group level phenomena (Alderisio et al 2017, Couzin 2018, Oullier and Kelso 2009, Riley et al 2011). Remarkably, the main empirical phenomena seen in the patterns of coordination produced by a single human being (except critical slowing down; cf Scholz et al 1987) have also been observed between people, such as the existence of two spontaneously stable coordination patterns (in-phase and anti-phase), transitions from anti-phase to in-phase at a critical movement frequency, critical fluctuations preceding the transition, and a shift in the phase relation between coordinating segments with different intrinsic frequencies (Kelso et al 1990, Schmidt et al 1990, Schmidt and Turvey 1994; see Schmidt and Richardson 2008, Tognoli et al 2020 for reviews). This means that synergies (and coordination in general) are grounded in informational coupling between intra- or inter-acting components or agents (Kelso 1994). The same patterns of coordination are observed regardless of whether the agents are body parts within a person (proprioceptive coupling); body parts and tactile, auditory, and visual stimuli (sensorimotor coupling); two people who watch, either intentionally or unintentionally each other's movements (visual coupling); humans and other species, as in riding a horse (which is strongly dependent on haptic coupling); and visually coupled humans and virtual partners or machines. In all these situations, coupling is by means of information exchange among parts and processes not by mechanical interactions. This does not mean that mechanical influences and neuromuscular-skeletal factors cannot play a significant coupling role, e.g., in stabilizing coordination (see Cuijpers 2019 for a nice example). But it does mean that fundamentally, coordination in general and synergies in particular deal in the currency of information that is specific to the dynamics (Kelso 1994, Kelso and Engstrøm 2006). For living things, synergies, as Sheets-Johnstone (1999/2011, 2015) says, are meaningful.
2. Understanding synergies: from synergetics to coordination dynamics
2.1. Basic forms of coordination; multiple modes; instability; phase transitions
Over 35 years ago Haken et al (1985) introduced a theoretical model (referred to in the literature as the HKB model) that has been adopted as a basic model system for understanding synergies. Originating from experimental findings (Kelso 1981, 1984) this model described (1) the coexistence of only two stable phase relations, in-phase and anti-phase (bistability); (2) the spontaneous switching between these coordinative modes (from anti-phase to in-phase); and (3) hysteresis in terms of the behavior of a multi-component system close to an instability, a so-called nonequilibrium phase transition. From synergetics (Haken 1983) it is known that close to instability points, where a system changes its behavior qualitatively, the number of relevant degrees of freedom is dramatically reduced and a low-dimensional description for the system can be found. The necessary information for a synergy to function is carried in collective variables or order parameters. The latter are relational quantities that are created by the cooperation among the individual parts of a system. They in turn may be seen to govern the behavior of the system's individual parts, in what has been termed 'circular' or 'reciprocal' causality. It is these functionally meaningful quantities and their dynamics that capture the qualitatively distinct patterns exhibited by complex systems as they unfold over time.
For all the cases of basic rhythmic forms of coordination within and between people the dynamics on a macroscopic level is captured by a single collective variable or order parameter: the relative phase between the interacting components. Haken, Kelso and Bunz postulated an equation of motion for the relative phase ϕ and derived it from a system of (nonlinearly) coupled nonlinear oscillators.
In the HKB model, the dynamics of the order parameter ϕ is captured by an equation of motion of the form:
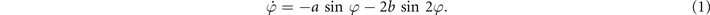
The properties of this equation are well-known and can be summarized as follows:
- For all values of a > 0 and b > 0 there are fixed points, corresponding to coordinated states at ϕ = 0 and ϕ = π
- In the parameter region b/a > 1/4 corresponding to low movement frequencies both fixed points are stable, i.e., movements can be performed either in the in-phase or anti-phase coordination mode
- In the parameter region b/a < 1/4 corresponding to high cycling frequencies, the fixed point at ϕ = π is unstable and the only remaining stable fixed point is at ϕ = 0 corresponding to an in-phase pattern.
The interacting components are given as

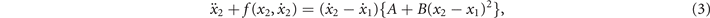
where each so-called 'hybrid' oscillator has the following functional form, again based on experimental evidence regarding the relationship between the amplitude, frequency and velocity of rhythmic movements:

The left-hand sides of equations (2) and (3) describe the motions of the individual components and the right hand sides the nonlinear coupling between them. The right-hand side of equation (4) consists of a linear self-excitation term,  , a Van der Pol term,
, a Van der Pol term,  , and a Rayleigh term,
, and a Rayleigh term,  , for saturation.
, for saturation.
Mathematical analysis shows that the relative phase obeys the dynamics of equation (1) with the coupling terms given as  and
and  where α and β are coupling parameters and R is the oscillator amplitude.
where α and β are coupling parameters and R is the oscillator amplitude.
The foregoing model system (equations (1) through (4)) reproduced the main experimental phenomena observed and predicted other properties such as increased relaxation times (critical slowing) and fluctuation enhancement as the transition is approached (see Fuchs 2013, Kelso 2009/2013 for reviews). There have been very many detailed mathematical developments of the so-called full HKB model ((1) to (4)), along with further experimental predictions and applications (for recent examples, see Cass and Hogan 2021, Leise and Cohen 2007, Slowiński et al 2018, 2020). The main focus here, however, is conceptual, and only the basic elements of the HKB style of modeling are included. These may be complemented, of course, by higher dimensional models of specific functions that may better match the complexity of the phenomena involved, e.g., in animal locomotion (e.g., Grillner 2006, Granatosky et al 2018, Holmes et al 2006; see also Schöner et al 1990) and disease states such as epilepsy (Houssaini et al 2020).
2.2. Fluctuations; symmetry breaking; self-organization
Variability is a crucial aspect of adaptive behavior in complex systems and plays a key role in exploring the space of possible coordination states as well as switching among them. The first extension of HKB therefore was to incorporate stochastic aspects (Schöner et al 1986). Now the potential corresponding to equation (1) takes the form:
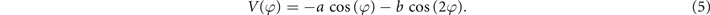
And the equation of motion is:
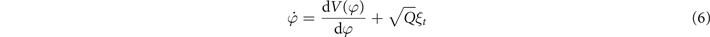
with  = 0;
= 0;  = δ(t − t') where Q is the strength of delta correlated Gaussian noise. Equation (6) was able to capture all experimental results on critical slowing down, switching time distributions and fluctuation enhancement with just a single free parameter. Notice in equation (5) symmetry plays a key role: the model system is symmetric with respect to both reflection (left and right are the same) and time (ϕ is 2π-periodic). Fluctuations incorporated in equation (6) are the source of spontaneous symmetry breaking. Why are fluctuations important? At the time of this work, the dominant theory was that movements were controlled and coordinated by homunculus-like motor programs stored in the central nervous system. Here, however, the introduction of the synergetic concepts of order parameters, stability, instability, phase transitions, etc and the intimate connection between theory and experiment led to the realization that coordination is a self-organized pattern forming process in which fluctuations and (in)stability play a critical role.
= δ(t − t') where Q is the strength of delta correlated Gaussian noise. Equation (6) was able to capture all experimental results on critical slowing down, switching time distributions and fluctuation enhancement with just a single free parameter. Notice in equation (5) symmetry plays a key role: the model system is symmetric with respect to both reflection (left and right are the same) and time (ϕ is 2π-periodic). Fluctuations incorporated in equation (6) are the source of spontaneous symmetry breaking. Why are fluctuations important? At the time of this work, the dominant theory was that movements were controlled and coordinated by homunculus-like motor programs stored in the central nervous system. Here, however, the introduction of the synergetic concepts of order parameters, stability, instability, phase transitions, etc and the intimate connection between theory and experiment led to the realization that coordination is a self-organized pattern forming process in which fluctuations and (in)stability play a critical role.
2.3. More on symmetry breaking; metastability; complementary pairs
Studies of coupling, not only between symmetrical body parts, but also between different body parts (an arm and a leg, for instance) or between environmental stimuli and limb movements led to a further development of the HKB model (Kelso et al 1990). In the latter case, syncopation—a key aspect of music, dance and drumming—corresponds to coordinating antiphase with a periodic stimulus, whereas in-phase corresponds to regular old synchronization. Remarkably, all the same phenomena observed in the original bimanual experiments (i.e., within the organism) occur also in the sensorimotor case (i.e., between organism and environment, action and perception)—along with several further phenomena that turn out to be important. Thus, in cases where the frequencies (ω1, ω2) of the coordinating parts are different, phase slippage and drift occur, intermixed with tendencies to coordinate near in-phase and anti-phase. This extended-HKB model (Kelso et al 1990, Fuchs and Kelso 1994, Fuchs et al 1996) takes the form:

where

The so-called metastable regime of the extended, symmetry breaking version of the HKB model opened up an entirely new way to conceptualize coordination in terms of complementary tendencies (Kelso 1995, 2012, Kelso and Engstrøm 2006, Tognoli and Kelso 2014). Thus, after the fixed points disappear, only remnants or ghosts of the previous attractors remain consisting of tendencies for the components to couple along with tendencies for them to remain separate. The metastable mode of operation is important for synergistic function: both the tendency for the parts (e.g., neurons, neural populations, etc) to come together and the tendency to express their individual autonomy coexist at the same time. The metastable mode provides synergies with a mechanism to generate far more flexible and fluid forms of coordination. In short, the foregoing interplay between experiment and theory indicates the synergy is: (i) a low-dimensional dynamical structure that is function- or task-specific; (ii) a cooperative phenomenon which exhibits both stable and flexible collective dynamics; (iii) the way nature simplifies control by reducing the number of degrees of freedom that need to be independently specified by 'control parameters' to accomplish a function; (iv) fundamentally metastable where the tendency toward togetherness (integration) co-exists with the tendency toward independence (segregation) (figure 1).

Figure 1. Bifurcation diagrams of the original HKB model (left, equation (1)) and the extended, broken symmetry version (right, equation (7)) with δω = 0 and 0.50, respectively. Lines correspond to stable fixed points or attractors qua coordinated states and dashes to unstable fixed points or repellors. On the left, the system is bistable, but at a critical value of k = b/a, the coordination state at antiphase disappears and switches to in-phase, where it remains. On the right, the same phenomena occur, but due to symmetry breaking the fixed points shift systematically before switching and eventually disappearing. Note that after the fixed points disappear, there is still attraction (phase concentration) to where the fixed points once were.
Download figure:
Standard image High-resolution image3. From the few to the many: generalized HKB model
The extended HKB model (equations (5) through (8)) has been demonstrated to explain bi- or multi-stability, phase transitions, metastability etc observed in many kinds of coordination in different biological systems and on different levels (Kelso 2021a for review). Although there have been elaborations to handle multi-component coordination, e.g. multilimb movements in humans and locomotory gaits in animals and insects (e.g., Golubitsky et al 1999, Granatosky et al 2018, Holmes et al 2006, Jeka and Kelso 1995, Righetti et al 2009, Schöner et al 1990), much of the research supporting the extended HKB model has involved the coordination of two interacting components, whether these be two joints of a single limb, two limbs working together, coordinating a single limb with discrete auditory, visual and tactile stimuli, two persons interacting with each other, humans coordinating with a virtual partner controlled by the HKB equations, and so forth (see Fuchs and Jirsa 2008, Tognoli et al 2020 for reviews).
An entirely independent approach aimed at capturing statistical features of large-scale coordination was developed by Kuramoto (1984) and became a paradigm for large scale synchronization in complex biological systems ranging from the flashing of fireflies, the firing of heart cells and neurons, to the awakened clapping of audiences composed of human beings. The fundamental idea of the Kuramoto model is that under certain conditions, a very large population of diverse elements undergoes a phase transition from incoherent, essentially random to highly coherent behavior in which the entire ensemble coordinates as a synchronized whole.
How might the large-scale dynamics of the Kuramoto model (equation (9)) be reconciled with the small-scale dynamics of extended HKB (equation (7))?

where ϕij = θ1 − θ2 and ωi corresponds to the dispersion of natural frequencies.
In the Kuramoto model the possible microlevel coordination patterns are difficult to study exhaustively due to the high dimensionality of the phase space. Although there are many mathematical analyses of the Kuramoto model in the literature (see Zhang et al 2019 for review), few if any have included empirical data on rhythmic coordination among multiple agents to justify their relevance to living systems. Experimentally, the large number of elements makes it difficult to perform systematic manipulations that scan the system's repertoire of possible patterns. Low-dimensional (macro) measures such as the overall level of synchronization may adequately capture the system's collective states but are insufficient to characterize coordinative complexity.
Recent experimental work on intermediate sized ensembles (in-between the few and the many) proves to be the key to uniting large- and small-scale theories of coordination (Zhang et al 2018). In this research, disorder-to-order transitions, multistability, order-to-order phase transitions, and especially metastability figure prominently on multiple levels of description, suggestive of a basic coordination dynamics that operates on all scales. The latter turns out to be a marriage of Kuramoto and HKB models (Zhang et al 2019).
The simplest model (equation (10)) that captures all important observations in the mid-scale human experiment of Zhang et al (2018) is a combination of equations (7) and (9).

Here φi is the phase of the ith oscillator, ωi the natural frequency, aij > 0 and bij > 0 govern the coupling strength.
This generalized HKB model (Kelso 2021a) handles how a diverse group of complex (human) agents interact with each other when their actions influence—and are influenced by—the rest of the ensemble in unpredictable ways and appears to be a key step toward embracing coordinative complexity. By going beyond synchrony, it promises a deeper understanding of collective behavior in all kinds of settings, from groups of neurons to groups of animals and people (e.g., Bardy et al 2020, Couzin 2018, Rio et al 2018, Warren 2018).
One nice aspect of equation (10) is that it allows the study of non-uniform coupling between agents and how interactions between relationships occur, e.g., in triadic situations (see Zhang et al 2019). Figure 2 is an illustrative example. The phase relationship between agents 1 and 3 has a dwell ∼ escape metastable dynamic: the relative phase pauses near zero (in-phase) at around 10 s and then drifts before returning again. Meanwhile the phase relationship between agents 3 and 4 hovers around zero, but bumps (shown by arrows) follow immediately after each dwell of agents 1 and 3. The longer agents 1 and 3 dwell with each other, the greater the perturbation effect on agents 3 and 4, a kind of critical slowing before they switch to out of phase with each other.

{kind=link}
Figure 2. Multiagent experiment of Zhang et al (2018) reveals interactions between agent relations. The evolving phase relationship (top) between agents 3 and 4 (ϕ34) is perturbed by agent 3's relationship to agent 1 (ϕ13) (see text for further explanation). Coordination patterns are captured by the orbits on an N-torus (bottom). The two relative phase patterns of interest are plotted from left to right on consecutive tori: rotation in the horizontal plane indicates change in ϕ13, rotation in the vertical plane indicates change in ϕ34. Notice the trajectory splitting in the two middle transitional patterns. Simulations (not shown here) indicate that a small change of the natural frequency of agent 3, can change the topology of the coordination from the leftmost to the rightmost picture.
Download figure:
Standard image High-resolution image{kind=link}
Visual interpretation of metastable coordination is easy when there are only a few agents interacting with each other. The situation becomes more complicated when there are more agents, hence more possible relations between agents, said relations evolving on multiple timescales. Traditional recurrence plot methods do not work in such situations and tools from persistent homology are needed. The resulting multiscale topological portraits prove to be revealing and suggest that transitions of collective patterns in complex systems of N interacting (human) agents are best captured by topology rather than states, per se. For example, in the case of complex dynamic interactions among eight agents, the recurrence of loops allows one to go back to the original data and identify global features that emerge spontaneously in time from local interactions (Zhang et al 2020).
4. Key concepts of coordination dynamics: order parameters/collective variables, intrinsic dynamics, and functional information
A key concept in coordination dynamics is that all the necessary information needed for the system to function is carried in the order parameters or collective variables. According to the Schöner–Kelso conjecture (cf Turvey 2004) a system's behavior is explained by identifying one or a few such relevant quantities that capture the qualitatively distinct patterns exhibited by the system as it unfolds over time and by describing these patterns (their emergence and dissolution) in the precise terms of attractors, bifurcations multistability, and related notions. The strategy of folding together all aspects within the dynamics of the order parameter/collective variable is an effort to embrace the full complexity of biological coordination without a proliferation of arbitrary divisions. In coordination dynamics, collective variables are deemed to span traditional distinctions between sensory and motor, perception and action, organism and environment, and dare one say, body (physics) and mind (psychology). In simple cases covered by HKB and related models, the relative phase, a dimensionless quantity, captures the coordinative relation between stimuli and responses in such a way that the latter predicts or anticipates the former. Much further work, however, will be needed to reveal the collective variables or order parameters that characterize coordination in more complex situations. In this respect, sophisticated measurement and analysis tools now available for studying creative patterns of coordination in natural situations and in real-time promise novel discoveries (see Balàgué et al 2013, 2017, Fuchs and Kelso 2018, Newell and Liu 2020, Passos et al 2009).
Intrinsic dynamics is equivalent to what Western medieval philosophy called adaequatio rei et intellectu, meaning 'the suitability of the cognitive equipment to its task'. For example, the concept of intrinsic dynamics operationalizes the pre-existing capabilities that a learner or learning system, at any point in time, brings into the learning environment, such as learning a novel task. The key conceptual aspect is that because the intrinsic dynamics (representing spontaneous pre-existing tendencies) is defined in the space of collective variables, any new information is defined in that space (Kelso 1995, p 162–163; see Kolchinsky and Wolpert 2018 for a formal approach). To grasp the idea, the great coach does not teach a behavioral pattern that lies outside the space of the performer's (or team's) intrinsic dynamics; he/she modifies the very variables that lie in that space. Indeed, knowing or intuiting the latter is what defines a great coach. The same argument can be applied at other levels of description (see below). Hence the term functional information: new information that modifies the (intrinsic) dynamics of collective, function-specific variables. According to coordination dynamics, if you want to change anything and have it persist 'permanently' (as opposed to being a mere transient) it is necessary to know the system's intrinsic dynamics. Knowing the latter means you know what to modify, and whether to use competitive or cooperative mechanisms to cause change. This principle operates at many levels, from individuals through society and may even apply to political change. Foreign policy, diplomacy and acts of aggression often flounder because of ignorance about the intrinsic dynamics of the system that they aim to influence or change. Obtaining measures of intrinsic dynamics in such situations constitutes a major challenge. The payoffs, however, are significant because it means you know what to change. Knowing the intrinsic dynamics is also at the core of so-called personalized medicine, for example in understanding why a drug has a positive effect on one person and no effect or a negative effect on another. Statistical studies in clinical populations hide this fact. Note again, the word 'individual' is not singular, but can refer to a group or a team or a tribe.
Though it is impossible to be inclusive here, it may be useful to highlight a few situations 2 where these concepts of coordination dynamics have been demonstrated to provide some insight and understanding, especially cases where theoretical models have been connected explicitly to experimental data...
- Learning. How new skills are learned and old ones modified, is a key issue for complex adaptive systems and a core determinant of cultural evolution (Waring and Wood 2021). In coordination dynamics learning is the process of modifying the pre-existing repertoire or intrinsic dynamics of the individual learner at neural, behavioral and social levels, captured typically as dynamical landscapes (Kostrubiec et al 2012, Nordham et al 2018, Zanone and Kelso 1992). In Kostrubiec et al (2012), changes in the dynamical landscape due to learning are shown to result from two basic mechanisms or 'routes': bifurcation and shift, the former representing qualitative, nonlinear change as a function of learning and the latter, smooth, linear change. Empirical data show that which mechanism is selected depends on the initial individual repertoire before the learning phase begins. Both bifurcation and shift mechanisms are accommodated by a relatively straightforward development of the HKB model. Model simulations show that the bifurcation mechanism results in greater stabilization of the to-be-learned behavior. Thus, stability not (or not only) error is demonstrated to be the basis of selection, both of a new pattern of behavior and the pathway of change that learning takes (smooth and linear versus abrupt and qualitative). Significantly, the dynamical feature of stability turns out to be a relevant feature upon which brain activity is organized while an individual performs a coordination task (for a nice example, see Jantzen et al 2009).
Recent work shows that teams or groups possess an intrinsic dynamics, captured as a manifold whose geometry and dimensionality are determined by task constraints and coordination requirements. Theoretically, the latter restrict the manifold to trajectories that are conducive to successful task performance (Dodel et al 2020; see also Pillai and Jirsa 2017, Tsuda 2018). Variability in the high-dimensional state space of neurobehavioral signals is seen to arise due to degeneracy, as noted earlier a fundamental property of complex biological systems that enhances flexibility and robustness (see, e.g., Greenspan 2012). Empirically, successful team performance is associated with dimensionality reduction of the manifold—evident, for example, in increased inter-brain phase coherence of beta and gamma rhythms during critical phases of task performance in which team members exchange information. The bottom line, and obviously an area for future research is that individual (within-brain) and social (between-brain) learning appear to be governed by the same dynamical principles (see Tognoli et al 2020 for review). Cooperative and competitive processes are intimately involved: the outcome of learning depends on whether new information cooperates or competes with the ongoing intrinsic dynamics.
- Recruitment and annihilation of degrees of freedom. One of the most characteristic though underrecognized features of complex biological systems is their ability to spontaneously (and simultaneously) recruit and annihilate whatever degrees of freedom are needed to accomplish goals and tasks. Patterns of coordination are assembled and disassembled with ease to accommodate functional demands. Whereas bifurcations within an already active set of components are now considered commonplace, e.g., in locomotory gait transitions where the ordering among limbs changes, the dynamical processes of flexibly recruiting and annihilating biomechanical degrees of freedom are only just beginning to be understood. Though often taken for granted, this is something that brains and organisms do effortlessly, but machines or robots do not. Again, using simple experimental paradigms as windows into this problem, dynamical mechanisms such as Hopf bifurcations reveal how, with changes in a single control parameter, fixed-points in one dimension can turn into limit cycles in two dimensions, and vice versa (Kelso et al 1993, Buchanan et al 1997). At the same time, theory and experiment have shown that biological systems can use the recruitment mechanism to freeze or stabilize coordination, under conditions in which patterns would otherwise destabilize and change (Fink et al 2000, Jirsa et al 2000). This is important because biological systems are both redundant and abundant (Latash 2008) and the simultaneous recruitment and annihilation of degrees of freedom is an important mechanism to both stabilize and destabilize coordination states.
- Brain dynamics. Coordination, it is sometimes said, achieves its pinnacle in the vast array of cells and connections called the human brain. A key feature of coordination dynamics is that it allows us to traverse (or more mundanely, connect) scales of organization from the behavioral level to the brain level through a combination of modern imaging technology, novel analysis tools and a neurobiologically realistic model of the cellular and neural ensemble properties of the cerebral cortex (see Kelso et al 2013 and Deco et al 2013 for reviews). Particularly relevant to physicists and complexity scientists is that the emergence of coherent spatiotemporal neural patterns with time-continuous dynamics may be described in terms of field variables. The connection between the brain level on one side and the behavioral level and environment on the other is achieved by embedding functional units in the brain (see Jirsa et al 1998; and Fuchs et al 2000 for specific examples). In such brain synergies, strongly inhomogeneous projections from one area to another afford directed information flow between spatially separate regions. Local changes in the connection topology can cause a global reorganization of neural activity (Jirsa and Kelso 2000). The connection topology may then be systematically used as a control parameter to guide the system through a bifurcation structure. Neural field models have continued to provide an overarching framework for connecting behavioral and neural dynamics, successfully handling coordinative effects observed experimentally, including, for example phase transitions in the human brain and neural correlates of motor equivalence (e.g., Kelso et al 1992, 1998, Mayville et al 1999). Moreover, they have been incorporated in a very promising modeling framework developed specifically for epilepsy and seizure dynamics called the Epileptor (Jirsa et al 2014, Houssaini et al 2020).
- Metastable mind? How are the multiple, diverse, and specialized areas of the brain coordinated to give rise to thinking and coherent goal-directed behavior? Active, dynamic processes like perceiving, attending, remembering and deciding—associated with the word 'thinking'—do not appear to be restricted to particular brain locations, but rather emerge as patterns of interaction in time among widely distributed neural ensembles, and in general between human beings and their worlds. A longstanding 'brain fact' is that neuronal assemblies in different parts of the brain oscillate at different frequencies (e.g., Basar et al 2001, Buzsàki and Draguhn 2004, Fries 2015, Kelso 1995). Historically, such oscillatory activity has been viewed as a possible instance of self-organization in the brain (Katchalsky et al 1974). According to coordination dynamics more crucial for cognition and action is the coupling or binding of brain rhythms together into a coherent network when individuals attend to a stimulus, perceive, remember, decide, act, etc, and when they interact with each other (Dumas et al 2011, Kelso 1995, Tognoli et al 2020, Valencia and Froese 2020). 'Not everyone', as two noted neuroscientists have recently remarked, 'agrees with the critical importance of brain oscillations but everyone acknowledges that neuronal activity should be coordinated across neurons and structures. Oscillatory rhythms appear the best candidate for this coordination role' (Buzsàki and Freeman 2015, p.v.). Poetry to the ears of coordination dynamics.
The extended, broken symmetry version of HKB promotes and predicts metastability (from meta meaning beyond) as a key dynamical mechanism for understanding how interacting components engage and disengage fluidly and synergistically over time (Kelso 1995). Metastable phase attraction between neural ensembles over multiple frequency bands has been proposed to explain how brains flexibly enter and exit coherent spatiotemporal patterns of neural activity (e.g., Aguilera et al 2016, Bressler and Kelso 2001, 2016, Fingelkurts and Fingelkurts 2004, Schwappach et al 2015). Fluid thinking, from the perspective of metastable coordination dynamics, is when brain rhythms are neither completely synchronized nor desynchronized. Instead of phase synchronized states that must be destabilized if switching is to occur, metastability consists of a subtle dwell and escape dynamic in which thinking is never quite stable and merely expresses the joint tendency for neural areas to synchronize together and to oscillate independently. Metastable coordination dynamics rationalizes William James (1890) beautiful metaphor of the stream of consciousness as the flight of a bird whose life journey consists of 'perchings' (phase gathering, integrative tendencies) and 'flights' (phase scattering, segregative tendencies). Both tendencies appear to be crucial: the former to summon and create thoughts; the latter to release brain regions to participate in other acts of being, knowing, and doing (Kelso 2008). Reducing the strong hierarchical coupling between the components of a complex system while allowing them to retain their individuality leads to a looser, more secure, more flexible form of functioning that promotes the creation of information. Too much autonomy of the component parts means no chance of them coordinating and communicating together. On the other hand, too much interdependence and the system gets stuck, global flexibility is lost. Well-known manifestations of too much synchronization in an individual brain, for example, are characteristic of diseases like Parkinson's disease and epilepsy.
Metastability is an entirely new conception of brain organization in which local segregative and global integrative processes coexist as a complementary pair, not as conflicting theories (Fingelkurts and Fingelkurts 2004 for review). As an expression of the full complexity of brains and people (Kelso 2001, Kelso and Tognoli 2017, Tognoli and Kelso 2014) metastability gives rise to a plethora of complementary pairs which may be said to constitute the complementary nature of coordination dynamics (Kelso and Engstrøm 2006). Complementary pairs like individual ∼ collective, integration ∼ segregation, competition ∼ cooperation, dwell ∼ escape, convergence ∼ divergence, states ∼ dispositions, etc fall out of metastable coordination dynamics. The tilde (∼) or squiggle symbol expresses a basic truth: both members of a complementary pair are required for understanding synergies and coordination. One without the other is incomplete. Metastability says that complementary aspects and their dynamics are found not just at the remote level of subatomic processes dealt with by quantum mechanics but also at the level of human beings, human brains and human behavior, that is, where the science of coordination plays out. Thinking narrowly in terms of contraries and the either/or is easy when life is simple. But in complex coordinated systems it seems that sharp dichotomies and contrarieties must be replaced with far more subtle and sophisticated complementarities.
5. Coordination dynamics of human ∼ machine interaction: virtual partner interaction (VPI) and the human dynamic clamp (HDC)
Unlike the laws of motion of physical bodies, the equations of coordination dynamics describe how the coordination states of a system evolve over time, as observed through special quantities called order parameters or collective variables. As we have noted, collective variables typically span the interaction of organism and environment. Imagine a machine whose behavior is based on the very equations that are supposed to govern coordination. Then imagine a human interacting with such a machine whereby the human can modify the behavior of the machine and the machine can modify the behavior of the human. The idea is analogous to the dynamic clamp used in cellular neuroscience to study the dynamics of interactions between neurons, but now scaled up to the level of behaving humans (a human dynamic clamp or HDC). This allows one to set up situations where the machine can cooperate or compete with the goal of the human and vice versa, with interesting (and unexpected) consequences such as the attribution of intention to the machine ('you are messing with me'). In the original virtual partner interaction (VPI) research (Kelso et al 2009), the embedded HKB model—on receiving input from human movement—drives an image of a moving hand which is displayed on a video screen. To complete the reciprocal coupling, the subject sees and coordinates with the moving image as if it were a real person observed through a video circuit. The key aspects of this social 'surrogate' are (i) that it can be precisely tuned and controlled—both by the experimenter and by the input from the human; (ii) that it allows for the study of mechanisms underlying real-time 'back and forth' information exchange; and (iii) that it provides opportunities to explore parameter ranges and perturbations that are not easily accessible in ordinary human interactions.
Whereas the initial VPI work examined quite simple forms of rhythmic coordination à la HKB, follow up research extended these to cover quite a broad repertoire of human behavior dealt with in coordination dynamics, such as models of both rhythmic and discrete movements, adaptation to changes of coordination with the environment, and learning, as specified by a 'virtual teacher' (Dumas et al 2014, Kostrubiec et al 2015). More recently, the human dynamic clamp was combined with high resolution electroencephalography, in order to identify the brain dynamics underlying self ∼ other relationships (Dumas et al 2020). Judgements of humanness of the HDC and measures of cooperation between human and HDC were found to modulate the functional connectivity between brain regions, particularly the right parietal region and the prefrontal cortex. This work opens up a way to understand how distributed neural dynamics integrates information from so-called 'low-level' sensorimotor coordination and 'high-level' functional information used for social cognition. Relatedly, the HDC was demonstrated to induce emotional reactions (as measured using skin potential responses) in human participants, especially when they believed the HDC was human and when coordination between human and HDC was stable (Zhang et al 2016).
A further practical, but rather worthwhile development is to use the HDC as a tool to quantify social abilities in children with autism spectrum disorder (ASD). In a group of 113 ASD participants, the HDC (here based on the HKB model) was shown to provide an accurate assessment of real-time social coordination, hence providing a clinical instrument to evaluate ASD and other neurodevelopmental disorders (Baillin et al 2020). For possible applications, this is just the tip of the iceberg: HKB coupling has been used to investigate social behavior between humans and small groups of simulated autonomous agents (Nalepka et al 2017, 2019) as well as provide a basis for situated robotics (Aguilera et al 2013, Cass and Hogan 2021 for recent analysis and review). The human dynamic clamp offers a path toward an intelligent, socially competent technological agent that will work and play with humans as if it were one of them (see Tognoli et al 2020 for review).
The way seems open to use tools like VPI/HDC to help rehabilitate and remediate brain disorders that impact social interaction, one of the cores of human existence. Were the HDC endowed with mathematical laws for brain (neural field) activity as well as behavior, one could subject it to lesions or neurological disease conditions. Physicists will already have noted that it is the symmetry between the human and the machine, and the fact that both carry the same laws of coordination dynamics, that is the essential aspect of the present approach. The design of the virtual partner mirrors the equations of coordination of the human neurobehavioral system. The laws obtained from accumulated studies describe how the parts of the human body, the brain and the environment self-organize, and address the issue of self-reference, a condition leading to complexity. It may even be speculated that the dynamically-based design of the human dynamic clamp qualifies as a sensorimotor equivalent of the turing test of humanness—whether a human judge can tell if an agent is a human being or a machine.
6. Directed self-organizing coordination dynamics
A main aspect of self-organizing dynamical systems is that the emergence of pattern and pattern switching occur spontaneously, solely because of the dynamics of the system: no specific ordering influence from the outside and no homunculus-like agent or program inside is responsible for the behavior observed. Yet somehow, that is, without magic or some vital force, what we call agency springs from the ground of spontaneous self-organized activity (Kelso 2002). Agency means action toward an end. How then do agency and goal-directedness come into the picture? A clue comes from studies of 3-month-old human babies (Rovee and Rovee 1969; see also van der Meer 1997). When babies are comfortable and lying in their crib, they kick their legs and move their arms spontaneously. After a while they tend to become fussy and so to amuse them parents will sometimes attach a mobile above their head that looks attractive and makes noises that babies seem to like. After a while, however, the baby gets bored or attention toward the mobile saturates. What if a ribbon is tied to the baby's ankles and attached to the mobile hanging over the baby's crib? By virtue of the coupling, any spontaneous foot or leg movements will cause the mobile to move. Under these conditions, using observational measures Rovee and Rovee (1969) reported that babies increase their kicking frequency by as much as a factor of 4. It is as if the infant realizes that it, not some outside force, is making the mobile move. When the baby perceives it is causing the world to change does it experience itself as an agent for the very first time? We are currently testing this idea in the laboratory on 15 full-term infants ranging in age between 2 and 4 months using a VICON motion analysis system which employs infrared cameras and retroreflective markers attached to the infant's body and to the mobile overhead. This allows for a 3D reconstruction of the trajectories of the infant and mobile markers in three-dimensional space sampled at 100 Hz (Sloan et al 2020).
Our hypothesis is that the key to the emergence of conscious agency is not only spontaneous movement but the bidirectional coupling (here haptic by means of the tether) between the baby and the world. Theoretically speaking, a functional synergy qua coupled dynamical system is formed when the (notably prelinguistic) infant discovers itself as an agent ('this is me'), that is, when the baby realizes it can make things happen. In this theory (Kelso 2016, Kelso and Fuchs 2016) the birth of agency and its causative powers ('I do', 'I can do') corresponds to a phase transition of a coordination dynamics whose key variables span the interaction between the organism (baby) and its environment (the moving mobile). The hypothesis is that the igniting of agency has a eureka-like, 'aha' effect; mathematically, it corresponds to a bifurcation in the coupled dynamics. Coupled dynamics refers to the coordinated relation or synergy between the baby's movements and the (kinesthetic, visual, auditory and emotional) consequences they produce. In our model (see Kelso and Fuchs 2016 for details) the main mechanism underlying the origin of self as a causal agent involves positive feedback between the end effector movements and the sensory effects they produce: when the baby's initially spontaneous movements cause the mobile to move, their perceived (or anticipated) consequences have a sudden and sustained amplifying effect on the baby's further actions. Thus, it is baby ∼ mobile (and in general subject∼object) coordination that is deemed to underpin the emergence of goal-directedness. This autocatalytic mechanism is continuous with our understanding of how biological form develops and of the feedforward network motifs so ubiquitous in the design of biological circuits (Alon 2007).
7. A theory of coordination: the main tenets of coordination dynamics
For Newton, 'the power of life and will by which animals move their bodies demonstrates there has to be yet undiscovered laws of motion'. For Schrödinger, 'the incontrovertible direct experience is that I am directing the motions of my body, of which I foresee the effects', this 'I' indicates (at least to Schrödinger) that 'new laws are to expected in the organism'. The Nobel physics laureate, Wilczek (2021) puts it even more bluntly: 'nobody walks, rides a bicycle, or catches a fly ball by working up from Newton's laws of motion, let alone the quantum theory of matter. To reach fundamental understanding, we need to rethink those experiences' (p 94). Here, by means of conclusions, we simply stake a (testable) claim for coordination dynamics, as the candidate set of ideas for a basic theory of biological coordination:
- (i)There are new laws of coordination in living things;
- (ii)Unlike the laws of matter and motion in inanimate matter, these laws describe the flow of coordination states produced by functional synergies or coordinative structures;
- (iii)Synergies or coordinative structures are the joint product of two forces (self-organization, followed by synergistic selection) and constitute the significant (relational) units of biological coordination at all levels;
- (iv)Coordination states on any given level of description are defined in terms of collective variables or order parameters that have been found to span traditional boundaries between organism and environment, stimuli and responses, perception and action, etc. Collective variables/order parameters and control parameters must be identified in complex systems;
- (v)The flow of coordination states produced by functional synergies/coordinative structures is written in the language of informationally coupled self-organizing dynamical systems, i.e., coordination dynamics;
- (vi)Multistability, phase transitions and metastability are prominent features of the coordination dynamics;
- (vii)The new laws of coordination apply to the coordination of very many things, just a few things and everything in between;
- (viii)Among other aspects, the laws address coordination within- and between bodies and brains, between organisms and their environments, between humans and machines, between humans and other species, etc. The laws are extensible and can handle adaptation and learning mainly through the concepts of intrinsic dynamics and functional information;
- (ix)What we call 'agency' or directedness emerges from the intra- or interaction of organism and environment spanned by collective variables/order parameters. The most primitive form of self-organization known in biological coordination (brains included), a synergetic phase transition, gives rise to the conscious self or 'I';
- (x)Metastability is an expression of the full complexity of brains and people and gives rise to a plethora of complementary pairs such as individual ∼ collective, competition ∼ cooperation, segregation ∼ integration, etc. Metastable coordination dynamics suggests that a deep principle of complementarity underlies life, brain, mind and society. The message of metastability is that in complex coordinated systems, thinking in terms of sharp dichotomies and contrarieties has to be replaced with far more subtle and sophisticated complementarities.
The foregoing statements are based on the results of over 40 years of empirical and theoretical research. They appear to be true as far as they go. Now, of course, the challenge is to extend and elaborate them for a deeper understanding of ourselves and the ever-changing world we live in.
Acknowledgments
Much of the material in this invited perspective is based on a plenary lecture entitled 'On Coordination' at the Conference on Complex Systems 2020 (CCS2020), held online Dec. 7–11, 2020. I am grateful to the Chairman, Prof. Panos Argyrakis and the Steering Committee of the Complex Systems Society for their kind invitation to present. The research described herein was supported by the NIMH, NSF, and ONR in the US, and the Human Frontiers Science Program abroad. I am grateful to all of them. Appreciation is also extended to the Davimos Family Endowment for Excellence in Science for their longstanding support. The research on infants mentioned in the text and the preparation of this article were supported by the FAU Foundation (Eminent Scholar in Science). This paper is dedicated to the memory of my long-term collaborator and colleague, the electrophysiologist and complexity scholar, Emmanuelle Tognoli.
Data availability statement
No new data were created or analysed in this study.
Footnotes
- 1
By and large, throughout this paper the reader is referred mostly to recent reviews which contain detailed references to background research studies.
- 2
It is impossible to cover all the basics in a brief perspective like this. A number of other situations of potential interest to readers, but not discussed here, for which experimentally supported mathematical modeling of coordination dynamics exist include: trajectory formation; anchoring; handedness; parametric stabilization; multifrequency, polyrhythmic coordination; multisensory integration; multilimb coordination; discrete and rhythmic behaviors; delayed feedback effects; cognitive influences, including perceptual, attentional and intentional aspects; social coordination and memory; human odometry, etc. Many of these phenomena have been studied using kinematic measures accompanied by fMRI, EEG and MEG to identify the corresponding brain dynamics. In addition, the concepts, methods and tools of coordination dynamics have been applied to a large range of skills such as ballet dancing, horse-riding, boxing, tennis, and team sports such as association football, rugby union, etc.