

Abstract
Climate change adaptation requires building agricultural system resilience to warmer, drier climates. Increasing temporal plant diversity through crop rotation diversification increases yields of some crops under drought, but its potential to enhance crop drought resistance and the underlying mechanisms remain unclear. We conducted a drought manipulation experiment using rainout shelters embedded within a 36-year crop rotation diversity and no-till experiment in a temperate climate and measured a suite of soil and crop developmental and eco-physiological traits in the field and laboratory. We show that diversifying maize-soybean rotations with small grain cereals and cover crops mitigated maize water stress at the leaf and canopy scales and reduced yield losses to drought by 17.1 ± 6.1%, while no-till did not affect maize drought resistance. Path analysis showed a strong correlation between soil organic matter and lower maize water stress despite no significant differences in soil organic matter between rotations or tillage treatments. This positive relationship between soil organic matter and maize water status was not mediated by higher soil water retention or infiltration as often hypothesized, nor differential depth of root water uptake as measured with stable isotopes, suggesting that other mechanisms are at play. Crop rotation diversification is an underappreciated drought management tool to adapt crop production to climate change through managing for soil organic matter.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Climate change will magnify the already persistent global challenge of crop yield loss to drought [1, 2]. Yield sensitivity and cereal production losses to drought have increased in recent decades, including for major global crops such as maize [3, 4]. Crop diversity has concurrently declined with trends toward simplification, specialization, and homogenization at both the farm and landscape scales [5] and such specialization has driven increasing sensitivity of agriculture to climate change in the U.S. Midwest [6].
Crop diversification and reduction in soil disturbance (tillage) could provide new opportunities to harness agroecosystem processes for resilient crop production [7]. Crop diversity lowers risk through the widely studied portfolio effect whereby different crops respond differentially to stress [8], but also possibly through emergent ecosystem properties like improved functioning and resilience, i.e. greater long-term stability due to higher resistance to abiotic or biotic stress and/or faster recovery [9]. Conservation agriculture approaches combining crop rotation, no-till, and crop residue retention increase rainfed crop yield in dry climates [10], implying potential to also protect crop yield from dry conditions in temperate climates. Long-term experimental sites including where this experiment took place show that diversification of temperate maize-based crop rotations increases yield, improves yield stability, and helps mitigate effects of weather variability on maize yield, particularly in hot and dry years and in no-till systems [11, 12]. However, the soil-plant mechanisms underlying greater crop drought resistance through adoption of these ecological intensification strategies have not been experimentally tested.
While improving drought resistance of agricultural systems has mostly focused on plant-centric strategies such as physiological differences between crop species or genotypes [13, 14], cropping system strategies considering the long-term impacts of agricultural management on soil properties and interactions with severity of plant water stress under drought have received little attention. Crop rotation diversification can impact multiple soil biological and physicochemical properties associated with building soil organic matter [15] and improving soil structure, water infiltration [16] and retention [17, 18] with potential benefits for yield stability [19, 20]. Improved soil structure can also affect root development and distribution by depth [21–24], which could facilitate root foraging and proliferation for water acquisition [25].
We empirically tested how crop rotation diversification increases maize drought resistance and potential underlying mechanisms. We imposed a seasonal drought along with ambient and supplemented rainfall controls in maize grown in plots with 36-year management legacies of crop rotation diversification and disturbance previously show in long-term data to span the range of maize yield in hot and dry years [11] in a major high-yielding maize-producing region. We measured a suite of soil and crop developmental and eco-physiological traits in the field and laboratory to test if maize plants are less water stressed under drought in diversified rotations and whether this relationship is mediated by shifts in soil organic matter and hydraulic properties as widely assumed [19, 20, 26] and/or by improved root development facilitating deeper or shallower plant water uptake [25].
2. Methods
2.1. Long-term crop rotation and tillage trial
The drought experiment was conducted at a long-term crop rotation and tillage trial established in 1980 at the University of Guelph Elora Research Station, Ontario, Canada (43° 38'' 27'' N, 80° 28'' 27'' W) [27]. The soil is a Woolwich silt loam, weakly to moderately calcareous Typic Hapludalf, and grey-brown Luvisol with approximately 4% organic matter and a 2% slope [27]. The site has a humid continental climate with 946 mm of precipitation annually and 88 mm monthly during the May–October growing season [28]. Average daily temperature ranges from 2 °C to 20 °C during the growing season, with cool springs and warm to hot summers [11].
The long-term trial is arranged as a randomized split-plot design with four replications and rotation as the main plot and tillage as the split plot (16.8 × 6.1 m) [11, 27, 29]. The drought experiment was conducted in first-year maize in three 4-year rotations along a rotation diversity gradient previously found to span the range of maize yield in hot and dry years [11]: maize-maize-soy-soy (MMSS), maize-maize-soy-wheat (MMSW), and maize-maize-oat/red clover-barley/red clover (MMOrcBrc). Maize, soy, wheat, oat, and barley were managed as cash crops and red clover was managed as a cover crop that was drill-seeded with oat or barley and grown in relay cropping after small grain harvest. Each rotation main plot was split into two randomly assigned levels of tillage: conventional tillage and no tillage. Conventional tillage consisted of moldboard plowing in the fall and secondary spring cultivation prior to crop seeding. Prior to initiation of no-till in 2002, no-till plots received reduced tillage (1980–2001) which comprised fall chisel plowing in most crops and the same secondary spring cultivation as in conventional tillage [11, 27, 29]. Fertility, pests, and weeds were managed conventionally to be non-limiting to crop yields in all phases of the rotation cycle as described previously for this long-term trial [30, 31]. During the drought experiment we report here all maize plots were fertilized with 11 kg N ha−1, 45 kg P ha−1, and 35 kg K ha−1 as 6–24-6 and 5–20-20 as a starter and 150 kg N ha−1 as UAN as a sidedress based on pre-sidedress soil nitrate testing. Crop cultivars for each crop changed over time to reflect increases in yield potential and typical practices of the region, with maize DKC 39–97 seeded 12 May 2016 at a rate of 78 000 seeds ha−1 and a row distance of 76.2 cm (30 inches) in this study period.
2.2. Manipulative drought experiment
In 2016, 36 years after long-term trial establishment, we modified the experimental design to a split-split-plot to accommodate a manipulative water experiment with three replications of each rotation-tillage-water treatment combination. Each rotation-tillage combination was split and semi-randomly assigned one of three levels of water inputs: drought, ambient rainfall, or ambient rainfall plus supplemental irrigation (supplementary information).
A seasonal drought was imposed in the field using below-canopy rainout shelters inserted between maize rows and elevated 30–40 cm above the soil surface on wooden stakes (figure S1 (available online at stacks.iop.org/ERL/16/084067/mmedia), supplementary information). All measurements were conducted in 4.8 m of the center row of the three maize rows where shelters were installed to avoid edge effects. The shelters were installed 22 d before the onset of maize anthesis and removed 35 d afterward to impose drought stress during flowering, the maize growth stage most vulnerable to drought [32] (figure S1). The percentage of rainfall intercepted by rainout shelters was estimated halfway through the drought imposition period by manually collecting rainfall that reached the soil surface both between plant rows and outside plots and calculating canopy throughfall and rainout shelter interception from water volumes. Irrigation volume in control plots was based on maize evapotranspiration estimates (supplementary information). A suite of crop developmental and ecophysiological traits and soil properties were measured in first-year maize for all rotations during the drought imposition period in 2016, except for infiltration and soil organic matter which were measured on the same rotation phase, first-year maize, in 2018 as noted in the following sections.
2.3. Plant growth, phenology, physiology, and yield
Maize phenology and growth were monitored during maize vegetative and reproductive growth stages as follows. Vegetative growth stage (leaf number) and height were measured 11–12 d after the onset of drought. Leaf Area Index (LAI) was measured 20 d after the onset of drought using a SS1 SunScan Canopy Analysis System (Delta-T Devices, Cambridge, United Kingdom). Maize shoot biomass was measured at 34–41 d after onset of drought and the end of the drought imposition. Maize shoot biomass was measured by destructively sampling three plants per plot, which were oven-dried at 80 °C for 2 d and weighed, then ground. For the latter date, biomass N concentration (% dry weight) was measured by combustion at SGS Canada Inc., Agriculture and Food (Guelph, ON, Canada).
We monitored three metrics of midday maize water status: xylem water potential (Scholander pressure chamber, Model 1000, PMS Instruments, Albany, OR, USA), stomatal conductance (gs) (SC-1 Leaf Porometer, METER Group, USA/Germany), and canopy temperature (FLIR thermal imaging camera T420bx), on three separate days during or within 2 d of maize anthesis (figure S1, supplementary information). Midday xylem water potential was measured using a Scholander pressure chamber (Model 1000, PMS Instruments, Albany, OR, USA). One shaded leaf per plot, at the height of the lowest non-senesced leaf in drought plots, was enclosed in a reflective aluminum-coated plastic bag for at least 60 min before removal from the plant and measurement. Stomatal conductance was measured using an SC-1 Leaf Porometer (METER Group, USA/Germany) on five plants per plot on a sunlight-acclimated, second-uppermost fully expanded leaf at midday. Canopy temperature at midday was estimated via whole-plot thermal imaging using a FLIR thermal imaging camera (T420bx) that was elevated by a pole 2 m above the crop canopy, centered on the sampling row, and captured its entire length excluding edges (4.8 m). Images were analyzed in FLIR Tools® software to quantify the average canopy temperature of the sampling row.
At physiological maturity, grain from all remaining plants per plot was hand-harvested, oven-dried at 80 °C for 2 d, and weighed. Reported grain yields were adjusted to 15.5% moisture content. Drought resistance was calculated as the relative (drought/irrigated) change in a given response variable for each crop rotation-tillage treatment replicate. Response ratios were converted to percent changes for data visualization. Less change due to drought indicates greater drought resistance.
2.4. Soil hydraulic properties
One undisturbed core per plot was taken at the end of the drought imposition in ambient rainfall plots, from a depth of 5–10 cm using 250 cm3 stainless steel rings to generate soil water retention curves via the simplified evaporation method (Hyprop, WP4 and Hyprop Fit software; METER Group, USA/Germany) (supplementary information). Water retained at field capacity (FC, −33 kPa) and permanent wilting point (PWP, −1500 kPa) was extracted from curves and available water capacity was calculated as the difference between FC and PWP. Bulk density was calculated as the dry soil weight per unit volume of each of the 250 cm3 undisturbed cores used for soil moisture retention analysis. Infiltration was estimated in 2018 in first-year maize in the same rotation-tillage treatment combinations as 2016 but different plots (each 4-year rotation sequence at the long-term trial is duplicated 2 years out of phase such that first-year maize occurs every 2 years). Field-saturated hydraulic conductivity (hereafter 'infiltration') was measured using an automated dual head SATURO infiltrometer (METER Group, USA/Germany) with one measurement per plot, which controls for variability in initial soil moisture by saturating the soil prior to measurement.
2.5. Soil moisture and organic matter
Prior to drought imposition, soil water potential sensors (MPS-2, METER Group, USA/Germany) were installed at 15 cm depth in one block (n = 6 per water treatment, one replicate of each rotation-tillage-water treatment combination), in order to monitor soil drydown under the drought treatment but not to characterize water treatment-rotation-tillage interactions. At 42–49 d after the onset of drought, one 4.4 cm diameter soil core per plot in the maize row was taken to a depth of 60 cm (hand sampler with slide hammer, Giddings Machine Company, Windsor, CO, USA) and divided into 10 cm increments (figure S1). Gravimetric water content was measured by weighing and oven-drying at 60 °C to a constant weight. In 2018, one soil core per plot in the maize row was taken in first-year maize in the same rotation-tillage treatment combinations as 2016 but different plots (each 4-year rotation sequence at the long-term trial is duplicated 2 years out of phase such that first-year maize occurs every 2 years). Cores (0–60 cm, 4.4 cm diameter) were divided into 20 cm increments (hand sampler with slide hammer, Giddings Machine Company, Windsor, CO, USA) and analyzed for soil organic matter (%) measured as percent loss on ignition at Ward Lab (Kearney, NE, USA).
2.6. Root distribution and stable isotope plant water sourcing
Roots length distribution by depth was quantified 42–49 d after the onset of drought using the same soil cores as for gravimetric water content (0–60 cm, 10 cm depth increments). Soil was washed from roots and roots were scanned and analyzed using WinRHIZO Pro software (Regent Instruments Inc., Canada) to quantify root length density (cm root cm−3 soil). Plant water sourcing from different soil depths was estimated 34–41 d after onset of drought using natural abundance of hydrogen stable isotopes in water extracted via cryogenic vacuum distillation [33] from simultaneously collected soil and plant samples and analyzed by mass spectrometry [34] (supplementary information). Hydrogen isotope ratios of the soil and plant water were used to assess plant water sourcing by soil depth using two methods. First, the estimated water uptake depth was identified by plotting isotopic compositions of each plant sample and average isotopic compositions of each soil depth within a rotation-tillage-water treatment combination in dual-isotope space and graphically identifying the soil depth with the most similar isotopic composition to each plant xylem sample [35]. Second, the proportional contribution of each soil depth to plant water was determined by averaging hydrogen isotopic composition for plant and soil depth replicates for each rotation-tillage-water treatment combination and generating full distributions of the possible proportional contribution of each soil depth to plant water using the stable isotope mixing model IsoSource with source increment set to 1% and tolerance to 0.05 [36].
2.7. Univariate statistical analysis
All statistical analyses were carried out in R 3.6.2. The lmer() command of the lmerTest package was used for linear mixed-effects model and the lm() command of the stats package was used in cases of linear fixed-effects model when all random effects accounted for zero variance. Analysis of variance (ANOVA) with fixed effects (rotation, tillage, water level, and soil depth where applicable), random effects (block, main plot, split-plot, and split-split plot where applicable) and all interaction terms was used to test the effect of treatments on plant traits and gravimetric water content. The fixed effect of water level, random effect of the split-split plot, and corresponding interactions were removed to test the effects of crop rotation and tillage on soil properties and drought resistance of plant traits. The assumptions of homogeneity of variance and normality of the residuals were assessed graphically using diagnostic plots and quantitatively. Response variables were transformed as necessary to meet assumptions. ANOVA was conducted using the anova() command of the stats package and means comparisons were conducted using the CLD() command of the emmeans package (version 1.4.3.01), both using Satterthwaite's method to approximate the degrees of freedom. In cases of multiple comparisons, the CLD() command used the Tukey method of p-value adjustment to compare a family of multiple estimates (alpha = 0.05).
2.8. Path analysis
We built a priori path analysis models based on current knowledge and hypotheses representing relationships between soil organic matter, soil hydraulic properties, depth of water uptake, plant water stress, and yield (figure S2). As visualized in figure S2, we expected increased organic matter to increase water retention at FC, PWP (a), and/or available water capacity (b), and/or water infiltration (saturated hydraulic conductivity), and/or shift the depth of root water uptake, through the effects of organic matter on soil structure and root proliferation [16–18, 25]. We expected that improved soil hydraulic properties and/or shifts in root water uptake depth would increase plant water status and yield. We then tested data fit to the a priori model using the lavaan package (v. 0.6–6). All data were standardized by centering and scaling to generate z-scores and a plant water status index was calculated (supplementary information). Following model fitting using the sem() command, non-significant relationships (p > 0.1) were iteratively removed and model fit was evaluated using the comparative fit index (CFI) (>0.95), root mean square error of approximation (RMSEA) (p < 0.05), standardized root mean square residual (SRMR) (p < 0.05), and Akaike's information criteria (AIC) [37, 38].
3. Results
3.1. Dynamics and severity of drought stress
The imposed drought spanned maize flowering by 1 month on either side. On average, the drought treatment using rainout shelters intercepted an estimated 56% of rainfall and reduced soil moisture matric potentials across the drought period by 670 kPa and 220 kPa compared to the irrigated and ambient rainfall controls, respectively (figures S1 and S3).
3.2. Crop rotation diversification maintains plant water status and yield under drought better than no-till
Crop rotation diversification improved maize yield drought resistance (better maintained yield under drought) (figure 1(a)) (rotation p = 0.037). Drought reduced maize yield by 53% relative to irrigated conditions in the lowest diversity rotation versus 36% in the most diverse rotation. No-till did not affect yield drought resistance (p = 0.678) nor rotation diversification benefits for yield drought resistance (rotation x tillage p = 0.737). Higher drought resistance with rotation diversification did not penalize productivity under ambient rainfall or supplemental irrigation conditions. Rotation diversification increased maize yield relative to the two-crop maize-soybean rotation across water levels, by 16%, 27%, and 56% under irrigated, ambient rainfall, and drought conditions, respectively (figure 1(b), rotation p < 0.001).
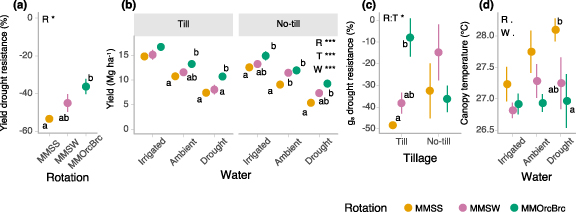
Figure 1. Effects of rotation, water level, and tillage on maize productivity and response to water stress: (a) drought resistance of maize grain yield, (b) maize grain yield at 15.5% moisture, (d) drought resistance of stomatal conductance (gs), and (d) canopy temperature. Drought resistance of maize grain yield and gs are defined as the drought-induced change in yield or gs as a percent of the irrigated control. Crop rotation abbreviations: MMSS = maize-maize-soy-soy, MMSW = maize-maize-soy-wheat, MMOrcBrc = maize-maize-oat/red clover-barley/red clover. Treatment factor abbreviations: R = rotation, W = water, T = tillage. Asterisks indicate significance of treatment effects from ANOVA (***p < 0.001, **p < 0.01, *p < 0.05, . < 0.1). Rotation means sharing the same letter or not lettered are not significantly different within a water and/or tillage level (alpha = 0.05). Error bars are standard error, n = 3 (b) and (c) or n = 6 (a), (d).
Download figure:
Standard image High-resolution imageCrop rotation diversification enhanced maize physiological drought resistance (better maintained plant water status under drought) (figures 1(c)–(d)) for two of three measures of plant water stress: stomatal conductance and canopy temperature. Maize in diversified rotations partially maintained stomatal conductance under drought relative to irrigated conditions in tilled systems (figure 1(c), rotation × tillage p = 0.026). In the two diversified rotations, the maize canopy maintained its temperature despite decreasing water inputs at temperatures similar to that of irrigated maize across all rotations. In the lowest diversity rotation, maize canopy temperature became progressively hotter (more stressed) as water inputs decreased (figure 1(d), rotation p = 0.073, water p = 0.085). Rotation diversification did not affect xylem water potential (whole-plant water status), which was higher (less stressed) with no-till but not with rotation diversification (rotation p = 0.830, tillage p = 0.018, water p < 0.001) (figure S4). Higher maize drought resistance with rotation diversification was observed despite these plants being taller with greater canopy development (higher LAI) (LAI rotation p = 0.030; height rotation p = 0.012, rotation × water p = 0.031, water × tillage p = 0.002) (figure S5(a) and (d)). Rotation diversification did not affect shoot biomass shortly after flowering but increased it at the end of the drought imposition period under drought (rotation p = 0.071, water p = 0.012, date p < 0.001). At 11–12 d after the onset of drought, maize vegetative growth stage was more advanced in the diversified rotation under drought and no-till ambient rainfall treatments but similar regardless of rotation diversification under irrigated and tilled ambient rainfall treatments (rotation p = 0.008, rotation × water p = 0.027) (figure S5(b)). No-till tended to reduce plant height (p = 0.051, tillage × water p = 0.002), leaf number (p < 0.001), LAI (p = 0.018), and plant population (p = 0.028) but did not affect shoot biomass (p = 0.438, figures S5 and S6). Rotation diversification improved shoot nitrogen concentration (water p < 0.001, rotation × water p = 0.043) while increasing maize shoot biomass, and better maintained shoot nitrogen concentration under drought (rotation p = 0.041) (figure S7), despite nitrogen application rates across all treatments being designed to be non-limiting to yield (see section 2).
3.3. Crop rotation diversification increases soil moisture with mixed effects on soil water retention and infiltration
Some soil properties that are often hypothesized to underlie plant drought resistance did not uniformly improve with crop rotation diversification. Soil organic matter (%) was highest in the topsoil (0–20 cm) (depth p < 0.001) and tended to increase with both rotation diversification and no-till but these effects were statistically nonsignificant (figure 2(a), rotation p = 0.146, tillage p = 0.238). Soil bulk density, an indicator of soil compaction, was affected by the interaction of rotation diversification and tillage (rotation × tillage p = 0.012), with lower bulk density (lower compaction) in diverse rotations in no-till systems but no effect of rotation diversification in tilled systems (figure 2(b)). Rotation diversification increased water retention at field capacity (rotation p = 0.080, figure 2(c)) with a similar trend at permanent wilting point (rotation p = 0.320, figure 2(d)). Available water capacity was affected by both rotation diversification and tillage (rotation × tillage p = 0.044) (figure 2(e)) and was highest in the most diverse rotations under tillage but did not vary between rotations under no-till. Field-saturated hydraulic conductivity, a measure of water infiltration, in the two diverse rotations was not significantly different from infiltration in the low diversity rotation, but was significantly higher in the intermediate diversity rotation than in the most diverse rotation (p = 0.020, figure 2(f)). Gravimetric water content 42–49 d after the onset of drought was interactively affected by rotation (p = 0.003), water treatment (p = 0.071), and soil depth (p = 0.035, water × depth p = 0.038) (figure 2(g)) and tended to increase with rotation diversification, though this effect was not statistically significant across all soil depths and treatment combinations.
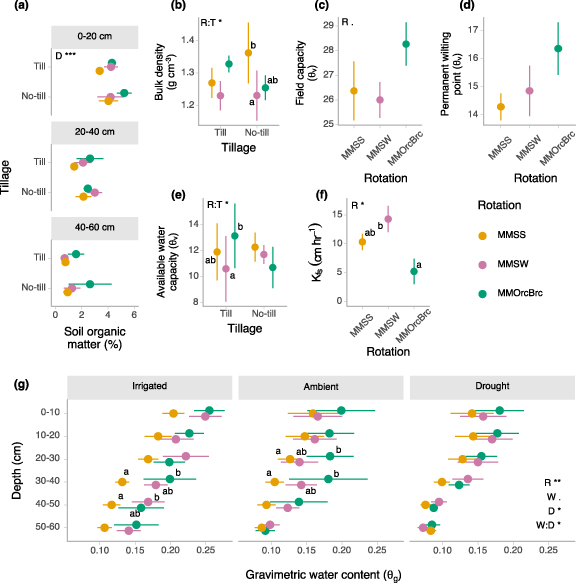
Figure 2. Effects of rotation and tillage on soil hydraulic properties: (a) soil organic matter, (b) bulk density (5–10 cm), water retained at (c) field capacity and (d) permanent wilting point (5–10 cm), (e) available water capacity (5–10 cm), and (f) field-saturated hydraulic conductivity. (g) Effect of rotation, tillage, and water level on gravimetric water content during maize grain filling (42–49 d after the onset of drought). Crop rotation abbreviations: MMSS = maize-maize-soy-soy, MMSW = maize-maize-soy-wheat, MMOrcBrc = maize-maize-oat/red clover-barley/red clover. Treatment factor abbreviations: R = rotation, W = water, T = tillage. Asterisks indicate significance of treatment effects from ANOVA (***p < 0.001, **p < 0.01, *p < 0.05, <0.1). Rotation means sharing the same letter or not lettered are not significantly different within a tillage level and/or depth (alpha = 0.05). Error bars are standard error, n = 3 (a), (b), (e) or n = 6 (c), (d) and (f), (g).
Download figure:
Standard image High-resolution image3.4. Tillage and drought impact root distribution and depth of water uptake
We found that root traits hypothesized to underlie plant drought resistance were not affected by crop rotation diversification. We assessed plant water sourcing by soil depth using natural abundance of hydrogen stable isotopes and sampled for root distribution by soil depth at 34–41 d and 42–49 d after onset of drought, respectively. The estimated depth of plant water uptake, the soil depth with the most similar isotopic composition to each plant xylem sample (see section 2), was most strongly affected by water treatment (p < 0.001): maize obtained water from increasingly deeper depths along a decreasing water gradient (figure 3(a)) and slightly deeper uptake depths in no-till (tillage p = 0.050). Full distributions of the possible proportional contribution of each soil depth to plant water from the stable isotope mixing model IsoSource show a similar pattern to main uptake depth results, with a greater contribution of shallow depths under irrigated conditions and a greater contribution of deeper soil layers under drought and ambient rainfall (figures S8 and S9). Water sourcing shifted from the topsoil to the subsoil under drought, but root length proliferated in the topsoil (figures 3(a) and (b)). Maize root length density (cm root cm−3 soil) was highest in the topsoil (0–10 cm) and was interactively affected by water treatment (p = 0.071), depth (p < 0.001, water × depth p < 0.001), and tillage (tillage × depth p = 0.096) (figure 3(b)). Root length density under no-till was more strongly concentrated in the 0–10 cm soil layer whereas under tillage root length density declined more gradually from the 0–10 cm to 20–30 cm soil layers (figure 3(b)).
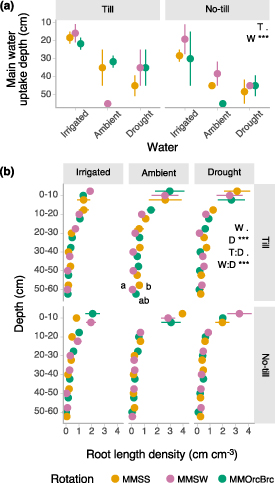
Figure 3. Effects of rotation, tillage, and water level on (a) estimated depth of plant water sourcing and (b) root length density by soil depth. The estimated depth of plant water uptake corresponds to the soil depth with the most similar isotopic composition to each plant xylem sample. Crop rotation abbreviations: MMSS = maize-maize-soy-soy, MMSW = maize-maize-soy-wheat, MMOrcBrc = maize-maize-oat/red clover-barley/red clover. Treatment factor abbreviations: R = rotation, W = water, T = tillage. Asterisks indicate significance of treatment effects from ANOVA (***p < 0.001, **p < 0.01, *p < 0.05, <0.1). Rotation means sharing the same letter or not lettered are not significantly different within a water and tillage level (alpha = 0.05). Error bars are standard error, n = 3 (n = 2 for panel a irrigated no-till MMOrcBrc).
Download figure:
Standard image High-resolution image3.5. Soil organic matter drives higher maize drought resistance but mechanisms are not fully resolved
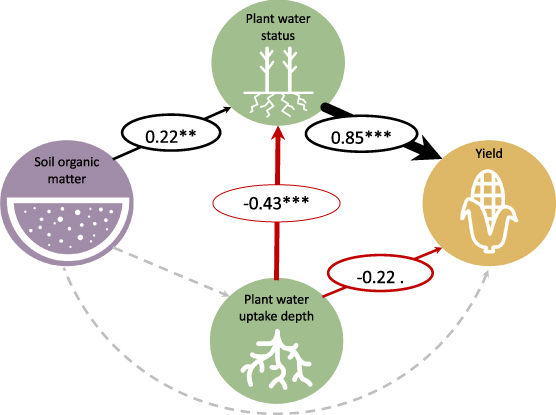
We found that soil organic matter strongly correlated with lower plant water stress and higher yield across treatments (figure 4). However, the positive relationship between soil organic matter and maize water status was not mediated the mechanisms of soil water provisioning or root water capture tested in this experiment. Soil hydraulic properties that are widely thought to underlie soil-mediated crop drought resistance (water retention at field capacity and/or permanent wilting point, available water capacity, infiltration) did not drive soil organic matter's benefit for plant water status, nor did shoot nitrogen concentration (path models with inferior fit and nonsignificant variables and paths not shown, see figure S2). An alternative mechanism tested, deeper water uptake by roots, was not associated with higher soil organic matter and correlated negatively with plant water status and yield.
{kind=link}
{kind=link}
{kind=link}
Figure 4. Final path analysis model of significant drivers of plant water stress, showing direct and indirect effects of soil organic matter on maize water stress and yield. Soil organic matter is in topsoil (0–20 cm); plant water status index is a function of stomatal conductance, xylem water potential, and canopy temperature measurements within one week of maize flowering, with better water status indicating less water stress; plant water uptake depth is the estimated soil depth from which the plant sources water; all data are z-scores (see section 2 for data processing and analysis). Standardized path coefficients are indicated for each significant relationship (asterisks indicate significance: ***p < 0.001, **p < 0.01, *p < 0.05, <0.1). Black, red, and grey arrows indicate positive, negative, and nonsignificant effects, respectively. Arrow widths are proportional to standardized coefficients. n = 49. Model fit criteria: SRMR = 0.008, RMSEA = 0.000–0.249 (90% confidence interval), CFI = 1.000.
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
Our results show that long-term diversification of aboveground plant communities through diversified crop rotations enhances crop performance and yield under drought likely through a belowground management legacy. We found that diversifying common maize-soybean rotations with small grain cereals and cover crops improves maize yield and physiological drought resistance and that soil organic matter is associated with lower maize water stress (figures 1 and 4). Our findings corroborate strong benefits of rotation diversification for maize in presumed and experimental drought years including in long-term data from the experimental site where this experiment took place [11, 12, 27, 39] with soil and crop physiological mechanistic evidence. This work advances crop diversification as a drought management tool not only at the farm or regional scale through a portfolio effect whereby different crops respond differently to stress [8, 40–42] but now at the plant-to-field-scale through improved soil-plant functioning and plant drought resistance. In the cooler temperate region where this experiment was conducted, rotation diversification was a more effective drought management tool than reducing soil disturbance with no-till, which did not impact drought resistance of maize yield (figure 1).
Crop rotation diversification may offer opportunity to both mitigate and adapt to climate change through investment in soil organic matter [43, 44], but linkages between soil organic matter and drought resistance (figure 4) are not fully resolved. Rotation diversification tended to increase soil organic matter (figure 2(a)) as found previously in meta-analysis [15] and consistent with trends in soil organic matter previously reported for these rotations at this trial [29, 31, 45–47]. We provide evidence that field-scale variation in soil organic matter, likely attributable to management, is associated with lower crop water stress, in agreement with global and regional trends that soil organic matter increases crop drought tolerance [48] and reduces crop sensitivity to precipitation variability [49] and yield losses and crop insurance payouts under drought [50]. However, rotation diversification and higher organic matter did not consistently increase water infiltration or retention (figures 2 and 4), in agreement with recent meta-analyses but contrary to our expectations and widely promoted benefits of soil health building management and organic matter for soil hydraulic properties [51, 52]. We also did not find evidence that deeper water capture by roots [25] or higher crop nitrogen status [53] mediate soil organic matter benefits for crop drought resistance (figures 3 and 4). Future research should test if soil biological mechanisms drive soil organic matter's benefit for plant water status, since changes in soil organic matter and biology are intrinsically linked [54] and shifts in the soil and rhizosphere microbial communities could play an underappreciated role in influencing crop drought resistance and productivity [55, 56]. Alternatively, increases in soil organic matter and biological activity may partially ameliorate negative effects of drought on nitrogen mineralization [53] or shifts in soil organic matter composition may play a role [47]. Reduced weed, insect pests, and disease pressure in diversified rotations [57–60] may also contribute to yield gains under drought, but their contribution is likely small at this site where weeds, insect pests, and disease are managed to be non-limiting to yield (see section 2).
The crop rotation diversification-driven drought resistance reported here underscores the need to reconcile breeding- and management-based approaches for building cropping system drought resilience. Amidst concerns of reaching a water ceiling for maize given higher temperature and evapotranspiration without gains in rainfall [61, 62], we highlight the underexploited potential of management-based approaches for improving plant drought resistance: rotation diversification increased maize yield by 56% under drought (figure 1(b)). Drought-tolerant maize hybrids increase yield by 5%–7% in water-limited environments, but the higher temperatures and atmospheric water demand (vapor pressure deficit) in these environments compared with our experimentally simulated drought precludes comparison of yield gains under drought through breeding versus rotation diversification [63, 64]. Crop models project increasing value of rotation diversification over time as yield potential increases in diverse rotations and soil organic matter declines in low diversity rotations under climate change scenarios, causing low diversity rotations to reach a water ceiling before diversified rotations [29]. Integrating management within breeding programs could provide synergistic benefits for climate change adaptation, reducing risks of crop failure while enhancing yield potential under high productivity and stress conditions [65].
Nearly 20% less maize yield lost to drought in diversified rotations provides a strong rationale to catalyze adoption and greater research and policy attention to help shield crop production from drought. That soil organic matter, rather that plant traits unique to maize, drives drought resistance suggests that rotation diversification may benefit drought resistance of other crops, consistent with evidence that rotation diversification increases soybean and small grain cereal yields in hot and dry years [11, 66, 67]. Although the mechanisms are not fully resolved, these results also highlight potential new levers for wider adoption of diverse rotations, such as integrating management and soil data including soil organic matter into crop yield risk and insurance models to improve the accuracy of crop insurance risk ratings [50, 68, 69]. Diverse rotations also have multiple co-benefits for agricultural sustainability including lowering synthetic nitrogen fertilizer and herbicide use [59, 70], insect pest resistance and damage [60], and freshwater toxicity [71], and boosting the disease suppressive potential of the soil microbiome [72].
5. Conclusion
We show that diversification of dominant, low diversity maize-soy rotations with common small grain cereals (i.e. wheat, oat, barley) and cover crops, enhances the climate change adaptation potential of a major global crop through soil organic matter, despite no significant increase in soil organic matter per se with diversification. Future research should zoom in, to understand microbial contributions to plant drought resistance and capitalize on the potential of biodiversity and plant-soil feedback for ecological intensification. It should also scale out, to further test the potential of crop rotation diversification to reduce yield risk to drought for maize and other crops and identify research and policy opportunities to incentivize rotation diversification for soil health and climate-smart agriculture.
Acknowledgments
Funding for this research was provided by the Ontario Ministry of Agriculture, Food and Rural Affairs and the Grain Farmers of Ontario. This work was supported by the Agriculture Experiment Station Projects no. CA-D-PLS-2332-H and the Foundation for Food and Agriculture Research. We thank the faculty, staff, and students who founded and have maintained and supported the long-term trial and acknowledge the particular contributions of Henk Wichers, Ramesh Eerpina, Hannah Toews, Michael March, Yixuan Wan, Rebekah Velasco, Vivian Tieu, and Cameron Ogilvie to trial management and data collection for the work reported here. We also thank Leonel Sternberg for hosting and training LR in the stable isotope component of this work. This material is based upon work supported by the National Science Foundation Graduate Research Fellowship Program under Grant No. 1650042 to LR. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation. LR's final work on this manuscript was carried out while funded by a Fondeyct postdoctoral grant (ANID Fondecyt de Postdoctorado 2021 N°3210036, Chile).
Data availability statement
The data that support the findings of this study are available upon reasonable request from the authors.
Author contributions
W D maintained the long-term experimental site where this study took place. W D and A G conceived and designed the experiment, with methods input from L R, M G, L S, and T M, and acquired the required funding. L R established the drought experiment and performed field sampling and laboratory analyses in consultation with A G and W D and analyzed the data in consultation with A G, W D, and T B. L R led writing the manuscript in close consultation with AG and it was critically revised by T B, M G, W D, L S, and T M. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare no competing financial interests.