


Abstract
As global climate change continues to impact regional water cycles, we may expect further shifts in water availability to forests that create challenges for certain species and biomes. Lowland deciduous riparian forests are particularly vulnerable because tree species cannot migrate out of the stream corridor, and they rely on root zone water availability that is controlled by variations in both local climate conditions (e.g. precipitation, evaporation, and infiltration) and non-local hydroclimatic forcing (e.g. streamflow, snowmelt, recharge). To determine how the seasonal water source usage of riparian trees is controlled by local versus non-local variability in hydroclimatic regime, we reconstructed the seasonal oxygen isotope (δ18O) signature of water used by two riparian tree species with contrasting rooting depths, comprising ∼800 δ18O tree-ring cellulose measurements from 12 tree-level decadal time-series at sub-annual resolution (six samples per year), along a strong hydroclimatic gradient within the Rhône River basin, SE France. These results were evaluated alongside δ18O measurements made from potential endmember water sources and independent hydroclimatic metrics. Thus we characterize the seasonal evolution of both potential water availability at distinct rooting depths and tree water source use and investigate the generalized riparian tree response to seasonal variations in local versus non-local hydroclimatic forcing over a decade. We show: (a) distinct seasonal water use between species, based on differential access to groundwater; (b) substantial source switching in both species based on evolving water availability; and (c) that riparian trees are more dependent on locally controlled soil moisture with distance downstream, creating increased vulnerability to locally increasing temperatures. We also find that deeply rooted trees in lowland riparian floodplains are potentially vulnerable to climate change because of their high dependence on water supply from mountains. This effect is more pronounced downstream, where seasonal water table decline may lead to loss of water required for deeply rooted trees.
Export citation and abstract BibTeX RIS
1. Introduction
Despite a growing consensus that forests across the globe are becoming increasingly vulnerable to water stress [1–5], insufficient information exists on the availability of specific water sources and the usage of this water by vegetation throughout the growing season (GS) across hydroclimatic gradients. Knowledge of water availability to plants and their use of this water in favorable hydrologic years versus drought periods will improve understanding of the proportional balance of groundwater versus infiltrated precipitation. It will particularly inform us about the evolution of key climatically controlled water sources for plant species rooted at different depths, and the corresponding vegetation responses to hydroclimatic forcing, whether locally or non-locally expressed. This knowledge gap is compounded by a lack of historical (i.e. decadal-multidecadal) baseline data, from which spatiotemporal trends of water source usage can be evaluated to predict and mitigate deleterious forest responses to shifting hydroclimate regimes under global climate change [1, 6, 7]. Lowland environments are of great concern because trees are already subjected to high levels of water stress, yet adaptation strategies such as upslope species migration are impossible because riparian trees in Mediterranean climates are adapted to lowlands and exist in isolated patches [8–12]. A new predictive understanding of the variation in water source availability and usage in response to local and non-local hydroclimatic forcing could support management interventions that prevent or mitigate large-scale forest diebacks, such as those recently attributed to increasing temperatures, precipitation variability, and root-zone moisture deficits [1, 7, 13–17].
Trees in riparian zones commonly utilize either phreatic (saturated) zone moisture or vadose (unsaturated) zone moisture sources, depending on their rooting architecture [18–21]. These water sources are comprised of an unknown mixture of regional groundwater flow, streamflow support of the shallow alluvial aquifer, and locally infiltrated precipitation [22–26]. Within seasonally dry climates (e.g. Mediterranean biomes), the cycles of the hydroclimate and vegetative growth may be out of phase, such that root zone soil moisture becomes limited during the summer period, when evaporative demand is highest and floodplain water tables are depressed due to declining streamflow. Small shifts in vegetative water stress resulting from changes in water availability could thus have important implications for tree growth and health, potentially driving shifts in forest community composition and function, ecosystem services, and carbon sequestration [27–31]. Climatic changes have already begun to significantly influence the water cycle in many seasonally dry regions, affecting both the input/output fluxes of water and the storage of water in soils, streams, and groundwater aquifers [23, 32–34]. Source water availability within riparian zones is also potentially affected by anthropogenic activities (e.g. dams, river training, groundwater abstraction), which are particularly common in Mediterranean systems, disrupting natural hydrological regimes and thereby impacting water availability at different rooting depths [22, 35–39]. The increasing likelihood of water cycle shifts in Mediterranean climate systems suggests the need for more detailed investigation of systematic patterns and timing of source water usage across known gradients of floodplain water availability.
One can conceive of water availability to vegetation in large river floodplains as a balance between (figure 1): (a) local climate, where shallow soil moisture is driven by the stochastic difference between precipitation inputs and evapotranspiration losses [40, 41], and (b) non-local climate, where water table elevation and corresponding capillary rise is driven by regional groundwater recharge and streamflow regimes that support the shallow phreatic aquifer [42, 43]. We refer to these as local and non-local hydroclimatic controls on water availability, respectively, noting that there may be marked differences in the magnitude, frequency, and timing of changes to the local and non-local water cycle that create differences in riparian zone water availability at any downstream location. Climate projections in Mediterranean-type ecosystems indicate that temperature and precipitation regimes will become progressively warmer and drier with less mountain snowpack [9, 44, 45]. These trends have the potential to alter the spatiotemporal availability of water within floodplains at different points along a river network, and thus to the overlying forest ecosystem [11, 46]. Nevertheless, our understanding of how hydroclimate is expressed in terms of seasonal tree water source availability is limited, which restricts our ability to adequately manage for the future consequences of climate change to Mediterranean riparian forests.
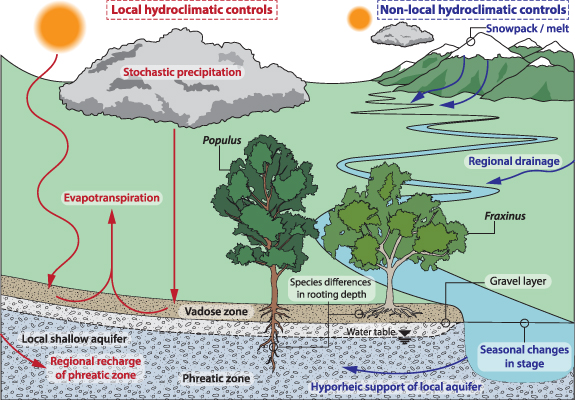
Figure 1. Schematic of local and non-local hydroclimatic controls affecting water availability to riparian trees and their use of available water.
Download figure:
Standard image High-resolution imageIsotopic ratios of oxygen (δ18 O) extracted from xylem samples, annual growth rings, and potential endmember waters have been particularly helpful in identifying and disentangling contributions from different source waters, where deeper water is usually associated with lighter isotopic values and soil water that has undergone evaporative losses becomes isotopically heavier [23–25, 47]. They have enabled improved understanding of the evolution of seasonally available water sources at critical periods of growth, and to identify the dominant water source usage over an entire season of growth [48, 49]. Recent developments in the study of δ18 O from tree ring cellulose have enabled the analysis of water source variability at sub-annual resolution in the same reference frame as the potential contributing endmember sources for co-occurring riparian tree species that typically use different water sources [48, 50]. This sub-annual information about water source usage is critical for predicting how riparian forests may respond to climatic changes that affect either local or non-local controls on seasonal water availability. Where there are substantial isotopic differences between river/phreatic water and precipitation/vadose zone water, δ18 O signatures support a relative comparison of source water uptake by riparian trees from shallow versus deeper floodplain water reservoirs, which represent local versus non-local hydroclimatic controls, respectively.
This paper aims to elucidate the dynamic behavior of hydroclimatic controls on subsurface hydrology through analysis of sub-annual patterns of tree water source use. We use seasonally resolved dendro-isotopic analyses of δ18 O extracted from tree ring cellulose, representing signatures of source waters (supplementary material), to generalize local and non-local hydroclimatic expressions of floodplain water availability. We conduct this study within three riparian forest sites spanning a strong hydroclimatic gradient along the Rhône River in SE France. These sites span a range of 3.5 °C and 140 mm in GS temperature and precipitation, respectively, over the period 2000–2010 from Peage de Rousillon—'Upstream' to Donzere-Mondragon—'Midstream' to Mas Thibert—'Downstream' (figure 2(b)). Along this hydroclimatic gradient, historical temperatures have already increased >2 °C between 1970 and 2015, yet precipitation trends are equivocal (figure S1 (available online at stacks.iop.org/ERL/16/084018/mmedia)). Each site contains common, and co-occurring Mediterranean tree species, Fraxinus excelsior and Populus nigra/alba (figure 2(a)). Fraxinus and Populus have been shown to exhibit contrasting rooting depths, where the former is restricted to soils above the gravel layer (∼ less than 1 m deep) and the latter has most of its root mass within gravels deeper than 1 m [31, 49]. The investigation utilizes an 11-year dataset of sub-annual tree-ring cellulose (δ18 Ocell) (n = 792) from individual, co-located Fraxinus–Populus pairs from both near channel (NC) and interior floodplain (IF) locations, within each study plot along the Rhône hydroclimatic gradient (figure 2(a)). We then employ an inverse biomechanistic model [50] to determine the δ18 O of source water (δ18 Omsw) utilized by each tree during the formation of each respective sub-annual δ18 Ocell sample (supplementary material, figure S2).

Figure 2. Map of the study region and field sites (a) with mean GS (May–September) temperature and total precipitation for each site (b). Numbered boxes correspond to the nearest climate stations for each field site: 1 = Bron, 2 = Montélimar, 3 = Arles Tour du Valat. Aerial imagery from GoogleEarth ©2017, CNES/Airbus, Maxar Technologies. Imagery dates: Upstream—22/08/2016, Midstream—07/9/2015, Downstream—26/03/2016.
Download figure:
Standard image High-resolution imageOur broader goal was to investigate how potential water variability along the hydroclimatic gradient affects water source usage by riparian trees. To accomplish this, we statistically compared distributions of δ18 O via Student's t and Mann–Whitney tests to identify site differences (and similarities) in potential water sources to riparian trees rooted at different positions. We then compared distributions of δ18 Omsw between tree species and along the hydroclimatic gradient via the Kolmogorov–Smirnov test, and we interpreted the median values in the context of δ18 O for these potential water sources—separately analyzed river, phreatic, vadose, non-GS (NGS) precipitation, and GS precipitation (supplementary material). We then use seasonal δ18 Omsw data for all growth years to develop characteristic patterns of seasonal water use for each tree and species along the gradient.
Finally, we evaluate these characteristic seasonal δ18 Omsw time series for each site against stream stage records (representing non-local controls on water table elevation) and a drought index (representing local controls on soil moisture availability as a function of the cumulative water balance over an antecedent period). Specifically, we assessed non-local controls on water availability in the shallow alluvial aquifer based on characteristic seasonal variations in upper percentiles of local stream stage within the Rhône River at each site, which integrates water production over the entire upstream drainage area. And we quantified water availability in the vadose zone based on the standardized precipitation and evaporation index (SPEI) [51], a drought index that reflects the local balance over previous periods between input precipitation and evapotranspiration that affect soil moisture. SPEI is calculated based on potential evapotranspiration from atmospheric variables rather than actual evapotranspiration. Therefore, a positive value of SPEI over the GS suggests that soil moisture should be available to plants at some point in the GS based on the time integral of rainfall minus PET over the antecedent months. Thus, we connect the characteristic seasonal plant water usage for these differentially rooted tree species along the hydroclimatic gradient to the local versus non-local controls on water availability in the rooting zones of the floodplain at different positions within the river basin.
2. Results
2.1. Modeled source water use from dendro-isotopes and potential endmember sources
Generally, we find systematically higher values of δ18 Omsw with downstream distance along the climate gradient (ensemble medians of −11‰, −9‰, −7‰, upstream to downstream), consistent with the downstream warming climate (figures 2, 3(a)–(c), figure S1 and table S1). However, there is substantial variability in δ18 Omsw between species, rooting locations, and sites (figure S3(a), table S1). For example, at NC locations, Fraxinus δ18 Omsw values are on average isotopically heavier than those of Populus, consistent with shallower water sources (figure 3, table S1). We also find that some individual trees use a large range of water sources, evidenced by a broad distribution with a muted peak (e.g. IF Populus at Midstream, figure 3(b)). Furthermore, there was notable within-site statistical similarity in δ18 Omsw distributions between Populus trees at both within-site rooting positions for both Upstream and Downstream sites, as well as between Fraxinus trees at Midstream (figures 3(a)–(c)).

Figure 3. (a)–(c) Kernel density plots of all sub-annual δ18 Omsw values calculated for each NC and IF Fraxinus and Populus at each of the study sites. (d)–(f) Potential endmember water source δ18 O characterization for each site. The NGS and GS precipitation δ18 O are denoted as NGS ppt and GS ppt, respectively.
Download figure:
Standard image High-resolution imageIn terms of endmember water sources, the δ18 O values in river samples along the climate gradient were statistically similar (mean value of −10.5‰, figures 3(d)–(f); Figure S3), suggesting a well-mixed pool of non-locally derived water in the Rhône. In contrast, δ18 O in locally controlled GS and NGS precipitation (ppt) vary along the hydroclimatic gradient, becoming isotopically heavier from north to south, e.g. −5‰ to −3‰ for GS ppt (and relatively heavier in the GS). This downstream heavier trend is partially mirrored in the vadose zone samples (rising from −10‰ to −8‰, north to south), though the inter-site differences are not statistically significant. Additionally, we find that the vadose zone moisture at all sites appears to derive from the lower half of NGS ppt inputs, associated with winter precipitation under colder conditions (figures 3(d)–(f)). At Upstream and Downstream sites, there is statistical similarity between the average values for river water and phreatic water (figure S3b), reflecting a tight connection between the Rhône River and its alluvial floodplain aquifer, compared with Midstream, where an isotopically heavier groundwater source from the Massif Central contributes to the alluvial aquifer [48]. Populus trees at each site and at all but one rooting location (Upstream IF, where both species exhibit isotopically light δ18 Omsw) tend to use isotopically lighter water than Fraxinus trees (figure 3, table S1). Generally, lighter values of δ18 Omsw used by Populus trees corresponds more closely to non-locally controlled phreatic water sourced from the Rhône alluvial aquifer than it does with locally controlled vadose zone water, yet the substantial variability over the decadal time period indicates hydroclimatically controlled seasonal variations in water sources for both Populus and Fraxinus trees (figure 3, table S2).
2.2. Seasonal patterns of water use
The subannual δ18 Omsw values averaged over all years (figures 4(a)–(l)) indicate the seasonal evolution of source water δ18 O utilized by each tree at each rooting position and site along the hydroclimatic gradient, based on more than a decade of sub-annual samples. There are notable patterns that emerge. For example, Populus trees at all three sites and rooting positions begin their seasonal growth using relatively isotopically heavy water, after which their water use becomes progressively lighter (non-locally controlled) until the last stages of latewood development, when isotopic enrichment toward heavier, locally controlled δ18 Omsw is evident (figures 3 and 4(a)–(c)). This suggests that Populus trees undergo seasonal water source switching from shallow (locally controlled) to deep (non-locally controlled) and back to shallow rooting depths (figures 3(d)–(f)). In contrast, Fraxinus trees do not exhibit systematic patterns across rooting positions and sites, although there appears to be consistent seasonal water usage between years for Fraxinus within each site. For example, Fraxinus at Upstream begin seasonal growth using the lightest δ18 Omsw (reflecting locally controlled NGS ppt, figure 3(d)), which then rises and plateaus during earlywood formation (reflective of local evaporative enrichment), before rising again such that the isotopically heaviest water is used at the end of the GS (LW3, figures 4(a) and (c)).

Figure 4. Modeled source water δ18 O (δ18 Omsw) for the sub-annual δ18 Ocell samples obtained from each tree for the 2000–2010 period at study sites, Upstream (a)–(d), Midstream (e)–(h), and Downstream (j)–(l). Blue and red lines correspond to the sub-annual δ18 Omsw means for Fraxinus and Populus, respectively. Grey shading represents ±1 SD.
Download figure:
Standard image High-resolution imageWe recognize one clear consistency across all trees at all sites: a dip in δ18 Omsw for initial latewood growth (LG1 or LG2) followed by an increase in δ18 Omsw at the end of the GS for all sites (LG3). This result suggests systematic source switching during latewood growth to a relatively shallow water source that has undergone significant evaporative enrichment (heavier isotopic value). This pattern is most dramatically observable for the NC Populus at Downstream (figure 4(j)). The spread of annual δ18 Omsw values is largest at Upstream, and smallest for Downstream (figures 3 and S3). This result suggests more interannual consistency in water use with distance downstream along the hydroclimatic gradient, where trees are more constrained in their use of variable moisture pools.
2.3. Climatic controls on seasonal water availability
The variations in δ18 Omsw between tree species within and between sites along the hydroclimatic gradient in SE France support further analysis of the local versus non-local hydroclimatic controls on water availability (e.g. via river stage and SPEI), which affect which water sources are used at certain periods of the GS. The results of these analyses are presented in figure 5, along with the mean seasonal patterns of δ18 Omsw. The Rhône River stage data show an evolving pattern with distance downstream. Due to contributions of snowmelt runoff from the Alps, stage (h) is generally highest at the onset of earlywood growth (EW1) at all sites (figures 5(d)–(f)), and then declines more with distance downstream (0.3–0.7 m for h95 , from north to south, for example), before slightly rebounding at the end of latewood growth (LW3). The main differences in h percentiles between sites is when the low point occurs and to what level it rebounds. For example, the trough in h at Upstream occurs during earlywood production, while it occurs during the latewood growth period for Midstream and Downstream. This is consistent with the expansion of drainage area with distance downstream that delays the annual flood wave in the Rhône [52], particularly that which is derived from French Alps snowmelt runoff (Downstream lies downstream of the Rhône's confluence with the Alps-draining Durance River). Additionally, the seasonal variation in h increases with distance downstream, reaching a range of >0.7 m for all three h percentiles at Downstream. High values of h relative to the average generally drive the filling of the shallow alluvial aquifer under the floodplain, which drains more slowly than it fills, leaving behind high non-locally controlled water availability for deeper rooting species [53]. In general, the stage values at Upstream and Midstream are ∼5–6 m below the surface elevation at tree rooting positions, although h is only 1–2 m below the surface at Downstream (figures 5(d)–(f)), suggesting greater access of aquifer water to trees at this site.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 5. Mean sub-annual δ18 Omsw for sampled trees at each site and floodplain location (a)–(c), corresponding GS river stage percentiles (d)–(f) and mean seasonal SPEI series for 1–6 month timescales (g)–(i).
Download figure:
Standard image High-resolution image{kind=link}
The SPEI values at each site show a similar pattern for 1–4 months of antecedent water balance (figures 5(g)–(i)). For example, the 1-month SPEI rises to its peak at the end of earlywood formation (EW3), and then essentially flatlines for the rest of the GS, whereas the 2-month SPEI peaks at LW1, the 3-month index peaks at LW2, and the 4-month SPEI peaks at LW3. This pattern suggests that the accumulated soil moisture throughout the GS is most available to trees in the final stages of both earlywood and latewood growth. The SPEI values with longer antecedence reflect soil moisture surplus derived from locally controlled NGS precipitation (which will be exhausted earlier), while the 1-month antecedence SPEI, for example, mainly reflects contributions from GS precipitation (also locally controlled).
2.4. Hydroclimatic patterns in seasonal water use
In the context of seasonal changes in h and SPEI, the characteristic patterns of seasonal water use for each tree indicate differences in the availability of deep versus shallow moisture sources throughout the GS and corresponding source switching by both species to maintain growth throughout the season. We summarize the water availability results by these metrics separately for the initiation of growth, the middle of the GS, and the end of growth.
At growth initiation (EW1) Rhône stream stage (h) is highest for all sites, yet this does not necessarily correspond to the use of non-locally controlled phreatic water in early stages of growth because Mediterranean trees will often only access this water source once the water table declines, leaving behind isotopically light capillary water in deeper soil layers [54, 55]. Thus, the starting values of δ18 Omsw values vary dramatically between sites (figures 5(a)–(c)), depending instead on the isotopic signature of readily available water in shallow soil layers from locally controlled NGS ppt, the volume of which is apparently higher upstream and declines downstream (figures 5(g)–(i)). The >5‰ range in starting δ18 Omsw values at Upstream suggest access from deep versus shallow root for Populus and Fraxinus trees, respectively, whereas all trees at the Midstream and the Downstream sites appear to use similar water to begin growth (except for the NC Fraxinus, figures 5(a)–(c)).
In the middle stages of growth, characteristic use of isotopically light (relatively) water for all trees suggests availability and use of water from deeper rooting depths. Lighter δ18 Omsw in mid-growth are associated with non-locally controlled phreatic or capillary water supported by the passage of Rhône River snowmelt runoff wave and/or the accumulation of deeper soil moisture by locally controlled NGS precipitation several months prior to growth (figures 2(d)–(f) and 4). The trough of δ18 Omsw occurs around LW1, and here there are similarities in water use signatures for all trees at all sites, except the NC Fraxinus at Midstream and both Fraxinus trees at Downstream (figures 5(a)–(c)), which are apparently rooted more shallowly such that they access a different (more enriched) moisture pool that reflects lower antecedent moisture [56] (figures 5(g)–(i)).
In the final stage of growth, there is consistent isotopic enrichment in locally controlled soil moisture, which is evident in the δ18 Omsw data for each of the Rhône sites, particularly for the more shallowly rooted Fraxinus (figures 5(a)–(c)). In fact, the seasonal patterns of water use suggest that both riparian tree species are adapted to take advantage of soil moisture at the end of the GS, even if the isotopic value of this enriched moisture pool varies substantially year to year (figure S4). At this stage, water may become very limited in both deep and shallow soil layers, such that trees are using decreasing pools of locally controlled soil moisture that may be briefly replenished by summer rains (GS ppt), which are subjected to high evaporation losses, producing high variability in end-of-season δ18 Omsw. It is also clearest from the LW3 δ18 Omsw values that the hydroclimate gradient exerts a strong control on source water availability and isotopic enrichment.
3. Discussion
Seasonal water source use variations throughout the growing period are an expression of adaptive plant responses to annual variations in water availability in phreatic or vadose zone stores. Thus, characteristic patterns of seasonal water usage may develop under a stationary climate, where climatic expression within hydrology regionally (non-locally) and locally are predictable. Clearly, there will be favorable and unfavorable years in terms of water availability in these phreatic versus vadose zone stores, but water source switching between shallow and deep roots would support healthy growth if both hydrologic stores are not negatively affected at the same time (e.g. a regional drought affecting upland snowpack and lowland rainfall). The analysis presented here shows a seasonal cycle of water availability and use by two co-located tree species with contrasting rooting depths that reflects the influence of: (a) non-local climatic controls on phreatic zone (and deeper soil layers) water availability and use in the middle of the GS, especially by the more deeply rooted Populus; (b) accumulation of soil moisture from locally controlled NGS precipitation primarily benefitting Fraxinus; (c) contributions from locally controlled GS precipitation to shallow zone water availability; and (d) locally high temperatures resulting in soil drying within shallow layers at the end of the GS (figures 2 and 4). The local and non-local hydroclimatic controls on riparian water availability and tree water use are summarized schematically in figure 1. This figure shows that non-local controls on water availability are dominated by snowmelt from the mountains and their contribution to streamflow selectively affects deeply rooting species, whereas the local balance between precipitation and evapotranspiration affects water availability to shallowly rooting species in riparian forests.
The climate is becoming warmer in SE France (a ∼2 °C increase in GS temperature between 1970 and 2015, figure S1) and in many seasonally dry environments around the world, where riparian trees are adapted to varying water availability by switching water sources between deeper and shallower rooting zones to support their growth and strong hydroclimatic gradients [23, 48, 57, 58]. Under nonstationary climatic forcing, the local and non-local water balance may become perturbed. Upland precipitation may shift in phase from snow to rain, leading to an increase in winter flooding but also to a smaller spring snowmelt pulse. Additionally, warmer upland spring temperatures may cause earlier/faster melting of remaining snowpack, shifting the timing/magnitude of the snowmelt flood pulse (figures 5(d)–(f)). These non-local effects may be exacerbated at riparian floodplain sites by locally controlled warmer lowland summer temperatures in the absence of precipitation changes (figure S1), which may result in higher soil evaporation, leading to faster drying and/or to rapid exhaustion of soil moisture at deeper depths in the profile. Furthermore, a non-local shift to more regional rainfall and less snowfall would support to a higher contribution of locally controlled NGS precipitation to summer soil moisture [41–43]. All these changes would affect the availability of moisture in both phreatic and vadose zone storage at key periods of tree growth [42, 43], but it is challenging to anticipate where and by how much.
The increasing vulnerability of riparian forests to drought stress suggests there is a necessity to improve the understanding of seasonal water source availability to trees. For example, systematic differences in co-located trees of different species suggest different water use strategies that may play out over seasonal timescales. Considering the Rhône hydroclimatic gradient, we show an increasing influence of (locally controlled) vadose zone water availability with distance downstream towards warmer climates, especially for Fraxinus. This suggests that within warmer, drier environments in the southern Rhône basin, source water availability to trees is responsive to local drought conditions, or even individual very dry seasons, leading to strong vulnerabilities in Fraxinus because their rooting architecture gives them fewer options for source switching. In contrast, Populus, a phreatophyte with a deeper rooting architecture, is less susceptible to negative effects of soil moisture drying, especially where the water table is quite shallow (at Midstream, figure 5(f)). Downstream experiences the highest availability of Rhône-sourced phreatic water of any site, supporting uptake and growth of deeply rooted trees especially over the latewood period. However, the high spring values of h at all three sites likely support an elevated water table and/or capillary water in deeper soil layers after water table recession, potentially leading to high water availability of isotopically light water to deep roots during latewood growth (figures 5(a)–(c)).
Along the entire hydroclimatic gradient, Populus trees consistently use shallow phreatic water to support mid-season growth (figures 5(a)–(c)), suggesting that their key water source is non-locally controlled. This means that Populus and Fraxinus trees along the Rhône are both responsive and vulnerable to distinct aspects of the hydroclimatic system (non-local controls on the shallow alluvial aquifer for Populus and local controls on soil moisture for Fraxinus, figure 1). We suspect this disconnect in controls on water availability to co-located riparian trees with distinct rooting depths is a common feature of many lowland river floodplains. Under stationary climatic conditions there should be (on average) sufficient floodplain water for both species, and each could manage a season of water limitation arising from an unfavorable hydrologic year via below-normal growth [31]. The progression of the GS also brings an associated isotopic enrichment due to evaporation and higher temperatures during precipitation events and increased evaporation, leading to higher values of δ18 O in soil moisture at LW3 than in EW3. In general, deeper water sources may be exhausted by August due to water table decline and depletion of hyporheic bank storage [48, 53], especially for the drier (most downstream sites). This creates conditions under which trees of both species access this enriched soil moisture to complete their annual rings at the end of the GS. However, given that riparian trees are transpiring throughout the GS [59], the true soil moisture availability will generally decline throughout the season, especially because GS precipitation is generally low in Mediterranean environments. This situation poses risks to both riparian species in years of unfavorable streamflow, when they will compete for a dramatically limited water source.
A nonstationary climate, on the other hand, may pose species-specific challenges, depending on whether it affects the water balance locally or non-locally. For example, if climatic changes affect upland snowpack/snowmelt, it is likely to limit phreatic zone water availability (non-locally controlled), especially at locations most distant from the mountains, such that phreatophyte species (e.g. Populus) may lose their buffer against locally warmer and drier conditions, and therefore become highly sensitive to soil moisture deficits [60]. In contrast, if local GS temperatures increase (which is already happening along the entire Rhône, figure S1) and/or local precipitation declines (more equivocal), soil moisture deficits will selectively impact shallowly rooted species (e.g. Fraxinus). It is also possible for both climatic changes (local and non-local) to occur simultaneously, negatively affecting both phreatic zone and vadose water availability (drought conditions) in lowland riparian forests. If this should occur, the inter-species competition for water will favor the survival of the more drought-tolerant species, Fraxinus [56], which has made more investment in shallow roots, and it would have important consequences for riparian forest composition along this and other river systems.
Since water deficits have been identified as the major drivers of forest die-back [1], alterations to forest species composition (e.g. hydroclimatic changes may favor the expansion of a particular species) may reduce biodiversity and ecosystem services. We have shown here that climate change may affect locally versus non-locally controlled water availability, leading to correspondingly different responses in Fraxinus and Populus trees along the Rhône hydroclimatic gradient. Ecologically, these spatially restricted lowland riparian forests have no adaptive escape hatch to ameliorate the effect of such changes [8–12]. These riparian forest patches are ecologically isolated with few opportunities to expand their range or even recruit on new floodplain surfaces, given their adaptation to lowlands and anthropogenic pressures in lowland floodplains [61–63]. Thus, lowland riparian forests, especially in Mediterranean climate regimes are 'sitting ducks' for climate change, due to their dependence on locally and non-locally controlled root zone water availability (figure 1) that will undergo changes with atmospheric warming and alterations to precipitation regimes. We suggest that characterization of seasonal water source availability and tree water use enables identification of water supply issues to riparian forests, and therefore allows for potential intervention to climate change at regional (water management) and local (forest management) scales.
Acknowledgments
Support for this work came from the U S National Science Foundation (BCS-1660490, EAR-1700555), the U S Department of Defense's Strategic Environmental Research Development Program (RC18-1006). It was also partly funded by Labex DRIIHM (ANR-11-LABX-0010) as part of the Observatoire Hommes-Milieux Vallée du Rhône (OHM VR) and performed within the framework of the EUR H2O'Lyon (ANR-17-EURE-0018) of Université de Lyon (UdL) through the 'Investissements d'Avenir' program operated by the French National Research Agency (ANR). The lab work was done based on an in-kind Natural Environmental Research Council Facility Grant via the British Geological Society's Stable Isotope Facility (Melanie Leng). We also thank various collaborators in France who assisted with access and equipment for field work. These include: Hervé Piégay, Bernard Pont, Jérémie Riquier, and Kristell Michel.
Data availability statement
All data that support the findings of this study are included within the article (and any supplementary files).
Author contributions
CIS and MBS designed research and wrote the paper. CIS performed research and analyzed data.
Conflicts of interest
The authors declare no conflicts of interest.