

Abstract
Although natural terrestrial ecosystems have sequestered ~25% of anthropogenic CO2 emissions, the long-term sustainability of this key ecosystem service is under question. Forests have traditionally been viewed as robust carbon (C) sinks; however, extreme heat-waves, drought and wildfire have increased tree mortality, particularly in widespread semi-arid regions, which account for ~41% of Earth's land surface. Using a set of modeling experiments, we show that California grasslands are a more resilient C sink than forests in response to 21st century changes in climate, with implications for designing climate-smart Cap and Trade offset policies. The resilience of grasslands to rising temperatures, drought and fire, coupled with the preferential banking of C to belowground sinks, helps to preserve sequestered terrestrial C and prevent it from re-entering the atmosphere. In contrast, California forests appear unable to cope with unmitigated global changes in the climate, switching from substantial C sinks to C sources by at least the mid-21st century. These results highlight the inherent risk of relying on forest C offsets in the absence of management interventions to avoid substantial fire-driven C emissions. On the other hand, since grassland environments, including tree-sparse rangelands, appear more capable of maintaining C sinks in 21st century, such ecosystems should be considered as an alternative C offset to climate-vulnerable forests. The further development of climate-smart approaches in California's carbon marketplace could serve as an example to offset programs around the world, particularly those expanding into widespread arid and semi-arid regions.
Export citation and abstract BibTeX RIS
Introduction
From the 1980s to present, about one-quarter of annual fossil fuel and land-use CO2 emissions have been sequestered by natural terrestrial ecosystems, helping to curtail both the pace and magnitude of global climate change (Ciais et al 2014). However, questions remain over the sustainability of this natural CO2 sink in the 21st century (Friedlingstein et al 2006, Ciais et al 2014). Of particular importance are growing incidences of extreme heat-waves, drought and wildfire in the planet's drier regions (Mediterranean and semi-arid regimes), which collectively account for ~41% of Earth's land surface and support ~30% of the world's human population (Safrie et al 2005). With unmitigated greenhouse gas emissions through 2100, the semi-arid regions of the planet are expected to become even drier (Collins et al 2013). Recent studies show that the trend and interannual variability of terrestrial CO2 uptake are dominated by the response of semi-arid vegetation to precipitation and temperature (Ahlström et al 2015). Moreover, tree mortality has been linked to drought and heat stress in dry climates, posing threats to sustained C storage by forests in such regions (Allen et al 2010, Allen et al 2015, Anderegg et al 2015, McDowell and Allen 2015). Further highlighting the importance of grasslands, past studies have shown the adaptive capacity of grassland ecosystems to extreme weather events (Vicente-Serrano et al 2013), consistent with evolution of grasses in favour of traits that confer drought-tolerance and adaptations to wildfire (Craine et al 2013).
Cap and Trade is the most widely used C pricing instrument designed to reduce greenhouse gases (GHGs), especially CO2. Its utility arises from its reputation as a 'free market' solution to the problem of climate change. While Cap and Trade policy allows the regulatory body to limit or 'cap' net CO2 emissions, the price of CO2 is determined by the individual entities or organizations invested in the market, generally with a given set of floor and ceiling prices (www.c2es.org/content/cap-and-trade-basics/). Industries and businesses can also be given the option to offset a fraction of their CO2 emissions by monetarily investing in GHG reducing technologies and natural sinks, such as forest conservation projects (www.arb.ca.gov/cc/capandtrade/capandtrade.htm). However, given the greater than 100 year residence-time of emitted CO2 in the Earth's atmosphere (Archer and Brovkin 2008), understanding the total length of time for which C remains sequestered in natural offsets is a critical determinant of realized emissions reductions and avoided climate change risks in the future. Here, we use a set of model-based experiments to demonstrate that grasslands are, over decades to centuries, more robust C sinks than forests in drought- and fire-prone regions like California, which are already experiencing substantial climate change impacts.
Our study focuses on California, USA, which is known for its drought and fire prone semi-arid ecosystems. Moreover, the Cap and Trade market of California is globally the fourth largest in terms of amount of emissions covered after the European Union, Republic of Korea and the Guangdong province of China, and second largest after the European Union in terms of total auction revenue (ICAP 2017). Under the current legislation, natural and chemical (i.e. short lived climate pollutants) investments can be used to offset between 4% and 8% of greenhouse gas emissions in California's Cap and Trade market. Many of the offsets include US forest C projects, ~40% of which are located within the boundaries of California (figure S1 available at stacks.iop.org/ERL/13/074027/mmedia) (Air Resources Board 2016). Like many semi-arid regions on Earth, California has experienced substantial drought and heat-waves over the past several decades, with concomitant increases in wildfire and tree mortality in the past several years. Recent surveys suggests that more than 100 million trees have died in the past couple of years following the severe and protracted drought of 2014−2015 (Diffenbaugh et al 2015). With a climate that is progressively hotter and drier (Riahi et al 2011), wildfires are becoming more widespread, damaging and aseasonal. Such changes have been statistically linked to increases in greenhouse gas emissions at the global scale, indicating that continued global climate changes will have a large effect on California's local environment (Diffenbaugh et al 2015). The California Air Resources Board inventory analysis suggests that the carbon drawdown capacity of Californian natural lands was reduced by 799 GgC over the period of 1990−2010 (www.arb.ca.gov/cc/inventory/data/data.htm). The resilience and durability of future terrestrial C storage has far-reaching implications for appraising the efficacy of greenhouse gas mitigation policies, like the Cap and Trade system, which in turn has ramifications on achieving the legislated mitigation targets.
We examine the hypothesis that climate change impacts will reduce the amount of carbon stored in forest vs. grassland vegetation in semi-arid environments. We perform several model-based experiments in California via a factorial design under the conditions of actual vegetation, forest-only vegetation and grassland-only vegetation in response to the following climate change projections into year 2100, in order of decreasing precipitation: (i) RCP 2.6 (RCP2.6) (van Vuuren et al 2006), IPCC's ambitious mitigation scenario, estimated to result in ~0.3−1.7 °C of global warming by 2100 (Collins et al 2013); (ii) RCP 8.5 (RCP8.5) (Riahi et al 2011), the IPCC's business as usual scenario, leading to a temperature increase of ~2.6−4.8 °C (Collins et al 2013); (iii) Periodic drought (Cycl. drt.), which approximates cyclical (e.g. El Nino/La Nina) precipitation patterns over a 10 year period (Cole et al 2002); and (iv) persistent drought (Perm. drt.), consistent with historical records for mega-droughts in the western US (Stine 1994). See Materials and methods for the definition of drought. While the model simulates wildfires (Thonicke et al 2001), land use change is not considered in our analysis. We thereby place an emphasis on natural ecosystems and their potential to sequester net CO2 in the absence of direct human management interventions. Apart from an analysis of statewide C totals, we also present our findings on a per-hectare basis, which provides a 'land C intensity' perspective that is generalizable. We use this approach to estimate CO2 (in C units) savings of natural offsets over the short (2025), medium (2050) and long term (2101).
Materials and methods
We use the dynamic global vegetation model (DGVM) LPJ-GUESS (Smith et al 2014) for our model-based experiments. This model includes 12 plant functional types (PFTs), which are groups of plant species classified together based on assumptions of homogeneity (e.g. in vertical and horizontal canopy structure) (Smith et al 2001). Consequently, a high degree of averaging of processes and structure is involved in order to inform the parameters which regulate the physiological processes. The 12 PFTs of LPJ-GUESS include temperate evergreen needleleaf trees endemic to northern California and the Sierra Nevada Mountains, and C3 grasses both naturalized and endemic throughout the state (Barbour and Major 1977). The model is 'dynamic', which means that the spatial distribution of PFTs is determined by the local environment (soil, light, climate) of the respective gridcell as opposed to prescribing PFTs a priori. When environmental properties are unsuitable for the growth of all 12 PFTs, the respective region is left barren, i.e. devoid of vegetation. PFT distributions are also determined by competition among PFTs, thus representing ecological succession. This depiction means that, with a changing climate, the distribution of PFTs (representing vegetation types) will also change under the conditions of a set of physiological and ecologically competitive rules, dynamically. Consequently, although the model samples from all 12 PFTs in a given grid-cell, the model-outcome, as demonstrated in table S2 (for years 2000−09), selects for PFTs (Primarily temperate needleleaf trees and C3 grasses) which are statistically best-suited to the environmental properties (climate, light, soil) in space and time. Wildfire occurrences in the model is dependent on fuel load and litter moisture, thereby combining the influence of climate and vegetation on the risk of fire. The amount of disturbance caused by a particular wildfire event is determined by both the length of fire season and PFT-dependent fire resistance properties (Thonicke et al 2001). The fire resistance for grasses is 0.5 while that of trees range from 0.1−0.3, making grasses more resistant to wildfires than trees, which is roughly consistent with field-observations since in the event of a wildfire, when compared to trees, a smaller fraction of the biomass of grass is damaged. Since the DGVM's default PFT parameters were based on global mean conditions, the drought sensitivity parameters of the tree and grass PFTs were specified for California, such that the DGVM's initial conditions conformed with MODIS satellite observations (Friedl et al 2010) (figure S3). For finer-scale observations, tuned parameters of the PFT 'Needleleaf Evergreen Tree' were set to the distribution of Sequoia sempervirens (California redwood) according to www.na.fs.fed.us/spfo/pubs/silvics_manual/Volume_1/sequoia/sempervirens.htm.
LPJ-GUESS does not include prescribed terrain data and thus depends on climate forcing data to simulate spatial heterogeneity of PFTs, which is essential for reproducing the higher altitude conditions of the Sierra Nevada mountains. We used climate observations from the Climatic Research Unit (CRU) (Jones and Harris 2013) at a spatial resolution of 0.5° Lat/Long. For future projections, we used the climate trends of the GCM (general circulation model) FGOALS-g2 (Li et al 2013) for two RCP scenarios described below, which exhibit the least bias for California winter precipitation (Caldwell 2010). Atmospheric CO2 concentrations of the same scenarios were also used to drive the model. Simulations for these scenarios were run at a spatial resolution of 0.5 degrees Lat/Long and restricted within the geopolitical boundaries of the state of California. Model simulations were carried out from 1901−2101 after being initialised with a 500 year 'spin-up' using detrended early-20th-century climate in order to build up vegetation and soil carbon and nitrogen pools to an approximate steady state (Smith et al 2014). Simulation results presented were however from close to present-day to the end of the century, i.e. 2015−2101.
In this paper, 'drought' refers to meteorological drought i.e. lack of precipitation. Because of the lack of consensus on the broad definition of 'drought', we defined drought to occur at precipitation levels less than 50% of RCP8.5 (Riahi et al 2011). 50% of normal precipitation has been found to impact agricultural, urban as well as industrial activities (Wilhite and Glantz 1985). Thus in Cycl. drt., there is reduced precipitation every 10 years while in Perm. drt. there is reduced precipitation for the entire duration of the model-experiment.
We focus on the standard metric of NBP (Net Biome Productivity), defined as the net production of organic matter after respiration and fire losses (Noble et al 2000) and analyze results for (a.) all vegetation-types present (ACTUAL); (b.) only grasses (GRASS-only); and (c.) only trees (TREE-only). The ACTUAL model experiment tracks realistic, present-day California vegetation responses to climate change, whereas the GRASS-only and TREE-only simulations reflect single land cover experiments against which maximal C storage potentials (hypothetical) of given ecosystem type can be analyzed.
Simulations of the GRASS-only experiments were performed by preventing the growth of all woody trees in the model, thereby eliminating competition from all woody tree PFTs from the beginning of 'spin-up' through 2100. This was accomplished by manipulating the parameter (of LPJ-GUESS (Smith et al 2014)) for the minimum 20 year coldest month mean temperature for survival (tcmin_surv) for all woody tree PFTs to an abnormally high value (100 °C). Since the actual temperature will always be lower than 100 °C, from the model's perspective it will then mean that the ambient temperature is lower than tcmin_surv or the minimum survivable temperature, effectively making tree growth impossible since technically it will be too 'cold' for the growth of tree PFTs. Even though there is no competition from tree PFTs in GRASS-only, the growth, productivity and spread of grasses were still constrained by the climate and atmospheric CO2 concentrations. The opposite was done for the TREE-only scenario, i.e. competition from grasses were eliminated. The consequent vegetation spatial distributions are shown in figure S2.

Figure 1. Retreat or expansion of grasslands or forests, i.e. changes in grass (A) and tree (B) fractions in response to 21st century climate changes. Blue indicates net expansion, red represents net contraction. Forests display widespread net retreat in all future climates except for the conditions associated with aggressive global greenhouse gas reductions (RCP 2.6). Grasslands show the capacity to geographically expand with climate change, with an elevational migration into the Sierra Nevada mountains in extreme drought conditions (3. and 4.)
Download figure:
Standard image High-resolution imageTable 1. Cumulative net biome productivity (NBP) from 2015–2101 for all ecosystem types in response to different climate change forecasts. Positive values reveal a net C sink, i.e. removal of CO2 from the atmosphere, while a negative values imply net C source, i.e. release of CO2 into the atmosphere.
| (a) NBP (cumulative) per hectare from 2015−2101 (Mg C ha−1) | |||
|---|---|---|---|
| ACTUAL | GRASS-only | TREE-only | |
| RCP2.6 | 4.1 | 1.0 | 5.0 |
| RCP8.5 | −3.5 | 2.3 | −2.5 |
| Cycl. drt. | −4.9 | 2.3 | −4.5 |
| Perm. drt. | −8.8 | −0.3 | −9.6 |
| (b) NBP (cumulative) California total from 2015−2101 (Tg C) | |||
| ACTUAL | GRASS-only | TREE-only | |
| RCP2.6 | 153.7 | 35.0 | 195.6 |
| RCP8.5 | −140.8 | 96.9 | −92.2 |
| Cycl. drt. | −193.8 | 96.6 | −172.5 |
| Perm. drt. | −355.3 | −13.3 | −383.3 |
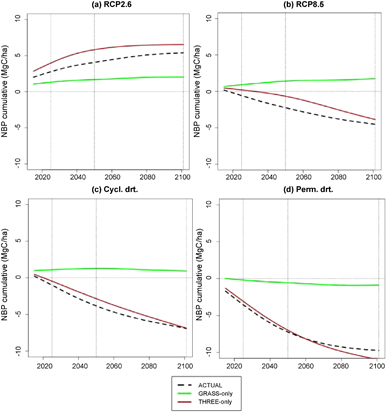
Figure 2. Net C storage change calculated as cumulative NBP after 2015 is used to compare the robustness of the respective ecosystems as net C sinks from the short to long term (shown by vertical lines). Positive NBP represents net C sequestration while negative values for net C emissions. The difference between the net C sequestration by grassland ecosystems increases progressively from the short-term to long-term. Results presented as smoothened lines for comparison (using a 'Spar' value of 1). Apart from the environmental factors and wildfires, vegetation shifts also impact NBP.
Download figure:
Standard image High-resolution image
{kind=link}
{kind=link}
Figure 3. Net C drawdown or loss in the 21st century, i.e. difference between the 2090s and 2010s (10 year means) of vegetation C (blue), litter C (red), and soil C (yellow) in response to 21st century climate change. Positive values reveal a net increase of a given C stock, negative values reveal a net decline of C. Units are in GgC (109gC). As drought, fire and heat intensify from RCP 8.5 (business as usual) to permanent drought scenarios, grasses emerge as the most reliable C sink. In the most extreme permanent drought conditions, grasses are a net C source but still maintain some C sink potential in litter and vegetation pools.
Download figure:
Standard image High-resolution image{kind=link}
The model LPJ-GUESS with the ACTUAL simulation experiment successfully captured the broad spatial distribution of California's vegetation (figure S3), the spatial pattern and magnitude of present GPP (Gross Primary Productivity) (figure S4) and soil C stocks (figure S5) vis-a-vis empirical observations statewide. It also simulated the winter greening and summer browning characteristic of a Mediterranean climate (figure S6). Furthermore, our simulated GPP falls within the range of multi-model GPP comparisons (figure S7). The modeled enhancement of productivity owing to the CO2 fertilization effect is consistent with free-air CO2 enrichment experiments (FACE) (Zaehle et al 2014). The ability of LPJ-GUESS to capture net carbon uptake responses to drought in the middle latitudes adds confidence to our prognostic modeling of future C storage in California (Ahlström et al 2012).
Results and discussion
In contrast to the conventional paradigm, we show that the inherent resilience of grassland vegetation to drought and wildfire (figure 1) translates to a more reliable C sink than forest ecosystems (figure 2) in response to 21st century climate changes. For all non-GHG mitigation scenarios, forests (TREE-only) store less C than grasslands (GRASS-only) across the ensemble of climate futures in California (table 1 and figure 2). The only case where forests are a more reliable C sink than grasslands is under the conditions of RCP 2.6 (van Vuuren et al 2006), which requires even more aggressive global greenhouse gas reductions than the Paris Climate Agreement. Indeed, the 'business as usual' climate change scenario (RCP 8.5) (Riahi et al 2011) points to grassland (GRASS-only) as the only viable net CO2 sink through 2101 (table S1 and figures 2 and 3). Grassland C storage is maintained under all conditions except for the most extreme mega-drought scenario, where soil C losses exceed C gains by vegetation and litter, thereby converting grassland ecosystems to a small but coherent net CO2 source (figure 3) through 2101. Differences in net C storage among grassland and forest ecosystems grow with drought intensity; the cumulative C sequestration of GRASS-only experiments exceed that of TREE-only simulations by 189.1, 269.1 and 370 TgC for RCP8.5, Cycl. drt., and Perm. drt., respectively, over the period of 2015−2101 (table 1, figure 3). The ability of grassland to mitigate risks of terrestrial C losses centers on the resilience of this biome to extreme heat, drought and wildfire, which become progressively more pronounced with increased greenhouse gas emissions (figures 2(a) and (b)) (Craine et al 2013 and Vicente-Serrano et al 2013).
The sustained grassland C sink is evident in both statewide projections and on a per area basis, pointing to both the geographic expansion of grasses into new areas and the ecological properties of grassland vegetation in maintaining future C sinks (e.g. Craine et al 2013). Despite boasting higher rates of GPP (figure S8), trees are unable to adapt to future climate changes in California. Compared to grasses, trees have a lower drought tolerance at low precipitation levels (i.e. 0−15 mm month−1), which reduces NPP compared to that of grasses (figures S11, a1 and b1).
Grassland vegetation, by contrast, dampens the magnitude of fluctuations in total C storage, with NBP oscillating around neutral values (figure S9, GRASS-only). TREE-only simulations show a much higher degree of NBP variability, switching from a strong net C sink to a strong net C source with the intensity of climate change in California. Our model predicts a more rapid gain of C per unit of precipitation under drier climates in grasses vs. trees (figure S11 b1). Grasses have evolved to thrive in semi-arid climates (Cherwin and Knapp 2012), developing physiological adaptations to overcome the challenges of drought-stress (Craine et al 2013) and protecting most of its C storage from fires by allocating it belowground. Furthermore, C allocation strategies in grassland vegetation allow for rapid resprouting from root-stocks, germination of seed banks, or both (Bond et al 2005, Bond and Keeley 2005, Nano and Clarke 2011) in response to wildfire.
The resilience of grassland vegetation to climate change is further highlighted in our simulations of ACTUAL vegetation in which the extent of California forests decline considerably more than grasslands across future climate change scenarios (figure 1 and S10). However, while the ecological 'banking' of grassland C to belowground stocks (roots and soil) helps to mitigate the risk of terrestrial C losses to the atmosphere, our simulations reveal a net loss of C from actual California vegetation (figure 3) in all scenarios other than RCP 2.6. This result points to the overarching risk of relying on traditional natural C offsets to reduce greenhouse gas emissions and highlights the need to consider changing environmental conditions while identifying the most reliable offset portfolio. It also implies that Cap and Trade markets should identify risky offsets and lower emission caps accordingly in order to avoid the risk of offsets switching from C sinks to C sources, which, in turn, can cause overall net emissions to rise.
Implications and caveats
Our study demonstrates that, in the absence of aggressive mitigation of global greenhouse gases, forest management strategies to reduce fire risks, or both, grasslands will store more C for a longer period of time than forests in California. Climate impacts create an environment which selects against trees in favor of grasses and other kinds of herbaceous vegetation. This result is consistent with empirical studies highlighting declining resilience of dryland forests to drought and wildfires (Stevens-Rumann et al 2017) and the adaptive capacity of grasslands to drought and fire (Craine et al 2013). However, the quantitative nature of our results are contingent on several simplifying assumptions, which should be considered when examining the implications of our work.
Generally, the GRASS-only and TREE-only scenarios are hypothetical single land cover scenarios used to contrast the resilience of grasslands and forests, respectively. The use of single land cover scenarios may exaggerate the magnitude of the difference in carbon drawdown potential between GRASS-only and TREE-only compared to real world where grasslands and forests likely co-exist. Moreover, since LPJ-GUESS allows environmental factors to determine the vegetation type in ACTUAL, area covered by GRASS-only and TREE-only do not add up to ACTUAL. TREE-only thus has significantly more barren land compared to ACTUAL (see table S2). Consequently, in the event of a wildfire, TREE-only has less carbon loss compared to ACTUAL. This explains why for the non-mitigation scenarios, the cumulative NBP (figure 2) of ACTUAL is lower than TREE-only.
Factors such as species traits, biodiversity, rapid evolution, and human management intervention could alter our model-based findings from the projections provided here. Consequently, our results indicate the potential direction of change as opposed to predictions that consider the full ensemble of ecological, physiological and management factors that can alter pathways and responses of ecosystems to climate change.
Specifically, LPJ-GUESS considers basic PFT mortality functions (Wramneby et al 2008), which fail to capture the 102 million dead trees that have been identified by US Forest Service's recent aerial survey (Stevens 2016). Indeed, testing the model against observations elsewhere have revealed the model's ability to only partially capture drought related mortality effects (Steinkamp and Hickler 2015). This means that trees may be even more vulnerable to drought-stress and fire than our simulations imply; though our model sensitivity analyses are consistent with higher drought and heat tolerance of grass vs. tree vegetation (figure S11.), consistent with expectations. On the other hand, the model also does not account for genetic plasticity, which makes grasslands better adapted to droughts and wildfires (Vicente-Serrano et al 2013, Craine et al 2013). This means that grasslands are more resilient in the real world compared to our simulations. Furthermore, our model simulates higher fire emissions (10.8 Tg C) over the period of 2006−2015 than statewide extrapolation of plot-measures in California (3.4 Tg C; (van der Werf et al 2010)). This difference could be due to issues of scaling, land use change, or fire suppression and fuel removal management practices, which are not included in LPJ-GUESS since it simulates 'potential' wildfire. LPJ-GUESS simulates wildfire as a function of fuel load and litter moisture, which, over regional to large scale studies, is consistent with observations (Abatzoglou and Williams 2016). Other important factors include herbivory/grazing and biodiversity, which have been shown to increase grassland C sequestration rates in data compilations (Tanentzap and Coomes 2012, Hungate et al 2017). Future work could focus on such factors as the evolutionary history of trees to fire, the physiological adaptations of ecosystems as well as regional species to fire, drought and climate change, the effects of biodiversity on ecosystem resilience as well as a comprehensive analysis of the goods and services provided by forests and grasslands.
With these caveats in mind, our findings have implications for understanding risks not only to carbon offsets in Cap and Trade programs but also to achieving greenhouse gas mitigation targets. Recent legislation has set California a target of reducing net GHG emissions to 40% of its 1990 levels by 2030. However the GHG emissions from potential wildfires as simulated by LPJ-GUESS from 1990−2015 ranged from 11%−50% of the 2030 target. These results imply further efforts on forest management interventions to reduce fire risk, and, given that tree-sparse grasslands are also working-land environments in California, including ranching, consideration of rangelands in Cap and Trade offsets could be explored for future offsets. While EU and California have championed market-based mechanisms such as Cap and Trade to reduce net greenhouse gas emissions, markets of China and Korea are in process of implementing their policies. Recent global and California focused studies (Griscom et al 2017, Cameron et al 2017) have emphasized forest conservation projects for emission offsets. Cap and Trade markets have thereby been structured to focus on aboveground C storage by trees as opposed to the longevity and resilience of net terrestrial C storage, especially in response to climate change. Hence the findings of this study could potentially serve as a lesson for other similar carbon markets around the globe. However, both EU and California's markets include forest conservation projects in developing tropical economies (e.g. REDD+ projects), which, on account of their higher moisture and productivity, may very well prove to be more robust C sinks than the forests of semi-arid environments, providing in addition, substantial co-benefits for biodiversity conservation, avoided land-use emissions and socio-economic justice.
In semi-arid environments, we suggest that C offset strategies should be designed in favor of long-term C gains to boost the reliability of Cap and Trade policies in meeting their emissions reductions goals. Incentivizing conservation of grasslands and rangeland practices that promote more reliable rates of C sequestration, for example, along the lines of the Marin Carbon Project (www.marincarbonproject.org/) may represent an important step toward conserving terrestrial C. Given that climate change impacts of CO2 emissions can last for centuries, explicit consideration of the vulnerability and longevity of terrestrial C offsets is urgently needed, with grasslands offering C sequestration services that appear to be more climate-adaptive in the rapidly changing and widespread semi-arid regions of the planet.
Acknowledgments
The National Science Foundation (grant EAR- 1411368) to BZH supported this research. The authors do not have any conflict of interest. Contribution by YPW is supported by the Australian National Environmental Science Program.