

Abstract
The Arctic Coastal Plain of Alaska is characterized by thermokarst lakes and drained lake basins, and the rate of coastal erosion has increased during the last half-century. Portions of the coast are <1 m above sea level for kilometers inland, and are underlain by ice-rich permafrost. Increased storm surges or terrestrial subsidence would therefore expand the area subject to marine inundation. Since 1976, the distribution of molting Black Brant (Branta bernicla nigricans) on the Arctic Coastal Plain has shifted from inland freshwater lakes to coastal marshes, such as those occupying the Smith River and Garry Creek estuaries. We hypothesized that the movement of geese from inland lakes was caused by an expansion of high quality goose forage in coastal areas. We examined the recent history of vegetation and geomorphological changes in coastal goose habitat by combining analysis of time series imagery between 1948 and 2010 with soil stratigraphy dated using bomb-curve radiocarbon. Time series of vertical imagery and in situ verification showed permafrost thaw and subsidence of polygonal tundra. Soil stratigraphy and dating within coastal estuaries showed that non-saline vegetation communities were buried by multiple sedimentation episodes between 1948 and 1995, accompanying a shift toward salt-tolerant vegetation. This sedimentation allowed high quality goose forage plants to expand, thus facilitating the shift in goose distribution. Declining sea ice and the increasing rate of terrestrial inundation, sedimentation, and subsidence in coastal estuaries of Alaska may portend a 'tipping point' whereby inland areas would be transformed into salt marshes.
Export citation and abstract BibTeX RIS

Content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Science has long recognized the presence of thresholds and instabilities within complex systems that can irreversibly transform them (Scheffer et al 2001). Some coastal segments of the Arctic Coastal Plain of Alaska constitute physical and ecological thresholds, scarcely elevated above sea level and extending kilometers inland. A small amount of terrestrial subsidence or sea level rise could inundate and alter these tundra areas. This potential is underscored by the presence of ice-rich permafrost, the surface of which may be vulnerable because of recent warming, particularly that associated with declining sea ice. Because the coastal plain is heavily used by migratory birds, caribou (Rangifer tarandus), denning polar bears (Ursus maritimus), and other mammals, the habitat transitions associated with such a shift could profoundly affect these species and the coastal terrestrial ecosystem.
Each summer thousands of geese migrate to the Arctic Coastal Plain to undergo their wing molt (Derksen et al 1982), during which time birds are flightless for approximately three weeks (Taylor 1995). This flightless constraint, combined with the energetic demands of feather synthesis, requires habitat characterized by high quality forage close to waterbodies used to escape predators (Madsen and Mortensen 1987). Aerial surveys conducted annually since 1976 by the US Fish and Wildlife Service demonstrate changes in mid-summer abundance or distributions of multiple species of geese within the Teshekpuk Lake Special Area (TLSA; Flint et al 2008). For example, the abundance of molting Black Brant (Branta bernicla nigricans, hereafter Brant) within the TLSA has remained stable, while the distribution has steadily shifted from inland lakes to coastal estuaries (Flint et al 2008). A recent study of molting Brant on several lakes across the TLSA suggested that improved habitat conditions along the coast likely explained the shift in goose distribution, but noted that further study would be required to identify the processes and extent to which habitat change had occurred (Lewis et al 2011).
Increased temperatures have affected Arctic coastal and near-coastal terrestrial environments through a variety of mechanisms. Shorter durations of sea ice (Serreze et al 2007), stronger storm surges (Arp et al 2010), and rising ocean and permafrost temperatures are among the causes of accelerated coastal erosion on the Arctic Coastal Plain north of Teshekpuk Lake since 1955 (Jones et al 2009, Jorgenson and Brown 2005). The accelerated nature of the coastline retreat suggests more frequent and extensive marine inundation of historically freshwater wetlands, particularly in these low-relief areas adjacent to inland waterways. Erosion and storm surge flooding have been suggested as the primary mechanisms responsible for land loss along the coast, though the processes responsible for altering coastal vegetation are unknown (Arp et al 2010).
The ice-rich permafrost soils and low relief of these Arctic coastal environments make vegetation highly sensitive to increases in high-water-mark or subsidence resulting from permafrost thaw. Saltwater inundation can kill typical tundra vegetation (Reimnitz and Maurer 1979), and repeated inundation likewise increases thermal conductivity, potentially triggering permafrost thaw and subsidence. Independently, terrestrial permafrost degradation and subsidence (Jorgenson et al 2006, Liu et al 2010) would also increase the area influenced by flooding from storm surges, snowmelt, or rain events. Sea ice loss is correlated with increased terrestrial temperatures (Bhatt et al 2010), including modeled increases in active layer depth (Lawrence et al 2008), but negative feedbacks associated with increased vegetation productivity could decrease thermal conductivity and negate these effects (Jorgenson et al 2010).
Our objectives in this study were to use time series imagery and soil stratigraphy to examine evidence of habitat alteration along the coastal zone north and east of Teshekpuk Lake that may have coincided with the observed shift in molting Brant. We hypothesized that flooding and permafrost dynamics were linked to habitat alteration, and that this alteration caused the observed shift in molting Brant distributions.
2. Study area and methods
We focused our study on two estuarine environments along the Beaufort Sea coast, Smith River and Garry Creek, both of which support increasing numbers of geese, as counted in July (figures 1 and 2; Flint et al 2008, Lewis et al 2011, Lewis et al 2010). Both estuaries are delimited by low-lying areas that are <1 m above sea level (figure 1; Mars and Houseknecht 2007), and are currently dominated by Carex subspathacea and Puccinellia phryganodes, which is classified as 'Halophytic Sedge (or Grass) Wet Meadow' (Viereck et al 1992) or 'Coastal Wet Sedge Tundra' and 'Coastal Barrens' (Jorgenson and Heiner 2003). Carex subspathacea exists exclusively as 'grazing lawns' in similar settings (e.g. Person et al 1998), where they are salt-tolerant communities (Person and Ruess 2003) inundated up to several times per year (Viereck et al 1992) and maintained by goose foraging ( Person et al 2003). These grazing lawns are comparable to those observed in our study area, though we did not determine the role of goose grazing in maintaining the vegetation. Both Carex subspathacea and Puccinellia phryganodes typically have high nitrogen content and are preferred forage for geese (Gadallah and Jefferies 1995, Person et al 1998, Person et al 2003). These halophytic meadows abut Lowland Moist or Wet Sedge Tundra (Jorgenson and Heiner 2003) at least a meter higher in elevation, which is dominated by sedges and underlain by characteristic ice wedge polygonal ground and ice-rich permafrost (Kanevskiy et al 2013). The active layer is typically 20–80 cm deep and permafrost ice content in these areas averages 73% underneath deltas and tidal flats to approximately 83% underneath the coastal plain surface and drained lake basins (Kanevskiy et al 2013).

Figure 1. Location of the Smith River and Garry Creek estuaries on the Arctic Coastal Plain of Alaska as seen by color-infrared imagery in 2002. Locations of soil pits are indicated (alpha-numeric), as are selected elevations (meters, in red; Mars and Houseknecht 2007), currents (rust) within the estuary, the flood prone zone (gray dash) and locations of other figures in the letter.
Download figure:
Standard image High-resolution image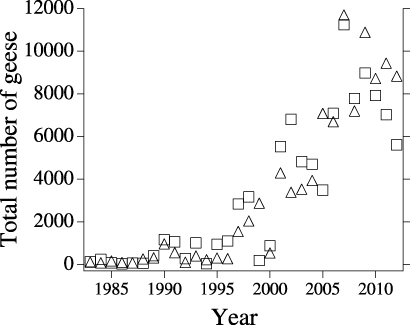
Figure 2. Increasing numbers of geese counted in July at the Smith River (squares) and Garry Creek (triangles) estuaries since 1983. Species include Black Brant, Greater White-Fronted Goose (Anser albifrons), Snow Goose (Chen caerulescens), and Canada Goose (Branta canadensis). Unpublished data from US Fish & Wildlife Service, Anchorage, Alaska.
Download figure:
Standard image High-resolution imageThe Smith River and Garry Creek originate near oriented thaw lakes approximately 18 and 10 km from their respective mouths in the Beaufort Sea. The only period of substantial freshwater flow in these drainages is during the snowmelt flood, when lowland areas are submerged for several weeks in May and June (Jorgenson and Heiner 2003, Meixell 2010–2013). Near their midpoints, both drainages transition from small low-gradient meandering rivers to amorphous estuaries. Astronomical tides are 40 cm (NOAA Prudhoe Bay tide station), and meteorological tides are regularly higher, with a century-level flood event triggered by a 1970 storm that rose approximately 2.5 m above sea level (Reimnitz and Maurer 1979). We consider Smith River and Garry Creek to be representative of the coastal portion of the TLSA, which has experienced the fastest retreat of any coastline between Barrow, Alaska and the Canadian border (Jorgenson and Brown 2005).
For the Smith River and Garry Creek drainages, black and white 9 × 9 in vertical aerial photographs from 1948 with sub-meter native resolution were scanned at higher resolution than the grain of the film; these were georeferenced in ArcMap 10.1 using >20 distributed tie-points per image and splines to modern 2.5-m scale satellite imagery. The georeferenced photos created contiguous coverage for the Smith River and Garry Creek estuaries. More recent imagery was obtained via USGS Earth Explorer (earthexplorer.usgs.gov) from the USGS mapping effort in 1955, and the Alaska high-altitude photography collection from the late 1970s and early 1980s; both sets were similarly georeferenced. A color-infrared aerial photo mosaic from 18 July 2002, and a SPOT mosaic from the growing season (green period) of 2009–2010, served as baseline imagery for georeferencing and for the qualitative landscape change detection. Considerable work has been focused on time series imagery chronicling the erosion of the Beaufort Sea coast (Jones et al 2009, Jorgenson and Brown 2005, Reimnitz et al 1988), but our primary interest was to assess changes to the salt marshes and adjacent tundra, so the higher resolution afforded by the 1948 photography permitted a longer and clarified assessment of landscape change, or lack thereof, in these areas.
During late July 2012, soil pits were dug to frost depth (25–68 cm) at eight sites along the 12-km length of the Smith River estuary, and five sites (only two shown) along a 1-km ocean to inland transect at Garry Creek (figure 1). To determine the history of goose habitat in these locations, all soil pits were dug in grazing lawns where goose feces indicated active use by geese. A soil block was extracted, cut with a knife to make a clean soil profile exposure, and then photographed next to a ruler. Soil horizons were sampled in 1 cm vertical increments and placed in 50 ml centrifuge tubes for transport to Fairbanks, Alaska, where they were frozen.
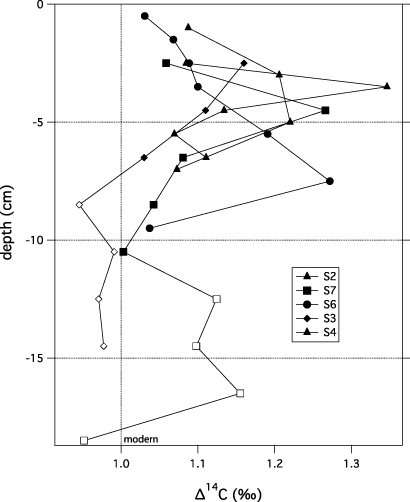
The University of California Irvine Keck Carbon Cycle Program used accelerator mass spectrometry (AMS) to radiocarbon (14C) date organic matter that we sampled from soil profiles (figure A.1). Bomb-curve and traditional 14C ages (n = 37) were calibrated to calendar AD dates using the Northern Hemisphere post-bomb calibration curve for post-1950 ages and the IntCal 08 curve for pre-1950 ages using OxCal 4.1 software (Ramsey 2009). At our sites peat accumulated vertically as organic material was added from plants fixing carbon at the surface. For plants that fixed carbon since 1950 we used 14C values of adjacent layers to establish ascending or descending slopes of the radiocarbon bomb-curve and assign calendar dates. The accuracy of 14C ages after 1955 is less than 1.3 year (Uno et al 2013), though the placement of the peak value from each pit on the ascending or descending slope caused uncertainty ordering on decades for some horizons. The 14C ages provided a timeline for soil stratigraphy; calendar dates assigned to above and below mineral layers were used to constrain the date of sediment deposition and changes in vegetation.
Vegetation assemblages from surface samples and buried vegetation strata were compared microscopically in pits S2 (0–2 cm, 4–6 cm), S3 (0–1 cm, 4–5 cm), S4 (0–1 cm, 6–7 cm), S6 (0–1 cm, 5–6 cm), S7 (0–1 cm, 4–5 cm), and G2 (0–1 cm, 3–4 cm). For each sample, ∼10 cm3 of sediment were washed and sieved through a 500 μm screen. What remained was examined under a dissecting scope at 10–50× power, and characteristic plant parts were isolated and photographed. Plant fragments were identified based on comparisons with scanned photos of herbarium specimens, taxonomic keys and illustrations of the potential species in the study area, and comparisons with plant fragments from known modern specimens collected at each sampling location. Nomenclature and habitats of all of the taxa follows Hulten (1968).
3. Results
The idealized soil stratigraphy from the Smith River and Garry Creek estuaries, created using the pit photographs and layer descriptions, shows Carex subspathacea growing in recent sedimentation overlying buried peat. Layered gray silt-rich sediment 4–7 cm thick, sometimes interbedded with vegetation horizons, overlaid peat >3 cm thick (figure 3). Underneath the peat was mineral soil or sediment. Microscopic examination of the buried peat showed that it was composed of predominantly undifferentiated graminoid remains. Samples from the surface vegetation and silt layer (0–1 cm) contained remains of Stellaria humifusa (Salt Marsh Starwort) in six out of six profiles, whereas it was not observed in any of the buried peat samples. Conversely, moss (identified to the family Amblystegiaceae) was absent from surface samples but common in buried peat, except at S7, where it was likely transported and deposited among the halophytic graminoids along with a Dryas leaf and worn wood fragment. In some locations the buried peat was underlain by a homogeneous gray unit (S1, S2, S6, G1, G2), while in other locations the buried peat was underlain by interbedded organic and sedimentary strata (S4, S5, S7). Coring the underlying permafrost revealed high ice-content (>60%) ataxitic cryostructure, comparable to what has been reported from tidal flats in this area (Kanevskiy et al 2013).

Figure 3. Soil pit stratigraphy and chronology at the Smith River and Garry Creek estuaries on the Arctic Coastal Plain of Alaska. Chronologies were derived from bomb-curve 14C at the Smith River estuary (top) and from time series imagery near Garry Creek estuary (bottom).
Download figure:
Standard image High-resolution imageAt Garry Creek the landscape changes associated with thawing of ice wedge polygonal tundra since 1948 were apparent at many locations in the time series imagery (figures 4 and 5), some of which we explored and verified in situ. Landscape changes in the Smith River area, except at the mouth, were subtle; the river channel and small ponds comprising the surface water in the estuary were consistently larger in the newer images. The type of vegetation in these areas and any change therein was seldom detectable in the imagery due to substantial differences between color perception recorded by the various photographic films and satellite sensors. Time series imagery from the Smith River and Garry Creek areas showed that coastal erosion occurred between 1948 and 1955, also altering the geometry at the mouth of the Smith River (figure 6); erosion thereafter is well-documented and has accelerated (Jones et al 2009).

Figure 4. Time series vertical imagery (left) and near-surface images (right) from Garry Creek on the Arctic Coastal Plain of Alaska. Images indicate permafrost thaw, subsidence, and sedimentation at soil pits G1 and G2 between 7/15/1948 and 7/17/1979 (left). The retreat of the coastline (dashed lines along coast) over the same period is small compared to the retreat of the inland subsidence front (dashed lines inland). A closer view of the subsidence front (upper right) shows lower elevations seaward, indicated by water pooling. A near-surface image looking seaward from the subsidence front (lower right) shows graminoids covering an ice wedge in the foreground. The brown background shows developing thermokarst pits filled with water among subsided salt-killed tundra and sediments being colonized by halophytic vegetation.
Download figure:
Standard image High-resolution image
Figure 5. Paired images near the mouth of Garry Creek on the Arctic Coastal Plain of Alaska from 7/17/1948 and 7/19/2002 showing thawing of ice wedge polygonal ground. Images (A) and (B) show the breaching of three larger ponds (squares) due to ice wedge degradation, apparent as the recent formation of lineal ponds overlying ice wedges (lower panels). Images (C) and (D) are near the mouth of Garry Creek and show a migrating subsidence front in polygonal ground (dashed black lines), leading to grazing lawn formation; during the same period, the coastline retreated negligibly. In the area between the dashed lines (D), subsidence is indicated by water in troughs of polygons.
Download figure:
Standard image High-resolution image
Figure 6. Composite of a 2010 SPOT image overlying a 1948 image, showing coastal erosion and changing geometry at the mouth of the Smith River, indicated by former shorelines derived from time series imagery.
Download figure:
Standard image High-resolution imageThe time series imagery and 14C chronology in soil pits suggests that an expansion of halophytic vegetation occurred during the last half-century. The 14C chronology indicates that the burial of peat and onset of sedimentation in the Smith River estuary occurred between 1957 and 1995, depending on the location, with an average date of 1973 (based on S2, S4, S6, S7; figure 3). The entire floodplain of the Smith River estuary, which extends approximately 8 km from the coast, has become a salt marsh in <50 years, during which time the coast at the mouth retreated approximately 1 km (figure 6). Soil pit stratigraphy indicates that the width of marine influence on vegetation in the Smith River area has increased from <1 km in 1957 (no halophytic vegetation at S2–S8) to 8 km in 2012 (figure 3), though the marine influence may have preexisted along the immediate margins of the river. At Garry Creek (soil pits G1 and G2), the aerial imagery indicates that this conversion occurred between 1948 and 1979 (figure 4). Near the mouth of Garry Creek, six areas where subsided polygonal tundra was evident in the imagery experienced an average three-fold increase (2002/1948; 537 m/296 m = 1.8; 205 m/45 m = 4.6; 411 m/191 m = 2.2; 200 m/28 m = 7.1; 376 m/370 m = 1.0; 295 m/192 m = 1.5) in the distance from the subsiding front of polygonal tundra to the shoreline (e.g. figures 4 and 5).
4. Discussion
These results provide evidence that vegetation has recently shifted toward halophytic goose grazing lawn in multiple areas near the coast of the Arctic Coastal Plain of Alaska. The combination of the 14C ages of the distinct soil horizons, the presence and absence of indicative plant remains, and the time series imagery show that, on average, vegetation change occurred in these areas in the 1970s (figure 3). We suspect that Carex subspathacea and Puccinellia phryganodes communities previously existed in a narrow band along the coast and inland margins, and that some combination of storm surge or snowmelt flooding, accelerated coastal erosion, sea ice decline and associated summer warming caused inland subsidence and sedimentation. Halophytic communities then expanded inland on the recently deposited sediment, and soil pits indicate that this vegetation change took at least a decade to spread throughout the Smith River estuary (figure 3). The timing of the vegetation changes described here fit with the observed shift in molting Brant distributions (Flint et al 2008), suggesting that the geese immigrated to new grazing lawns in response to an increase in extent of this preferred habitat. These findings provide support for the environmental change hypothesis put forth to explain the increase in abundance and novel distribution of four species of geese in the TLSA over the past 30 years (Flint et al 2008).
Profiles S1, S2, S6, G1, and G2 represent the clearest evidence of recent sedimentation, including a shift toward halophytic vegetation, apparently unprecedented for centuries at these locations. Profiles S3 and S8 also show a shift toward sedimentation, though not as clearly. Profiles S4, S5, and S7 exhibit the recent sedimentation and buried peat, but the units underlying the buried peat are similar, so it may be that this part of the estuary had active sedimentation prior to development of the vegetation layer, now peat. The presence of halophytes Carex subspathacea and Stellaria humifusa growing in recent sedimentation, and the absence of S. humifusa in the buried peat indicates a shift toward halophytic vegetation communities. The lack of moss in surface samples and abundance of moss in buried tundra layers also supports a shift toward halophytic vegetation (Jorgenson and Heiner 2003), implying novel saltwater inundation (Viereck et al 1992). The soil stratigraphy suggests that the shift is directional, not cyclical, because dates in the buried peat layer extend back to the 17th century (figure 3).
The new sedimentation and halophytic vegetation implies that either storm surge height has increased or permafrost thaw and subsidence of ice-rich permafrost has occurred. Evidence from Barrow meteorological records suggests a possible increase in storm surge flooding (Arp et al 2010), meanwhile permafrost thaw and subsidence is apparent at numerous locations near the mouth of Garry Creek where the landscape exhibits a gradation from elevated tundra polygons to subsided and sedimented grazing lawns (figures 4 and 5). What is unclear from our results is whether (1) storm surge or meltwater inundation preceded and caused subsidence, (2) permafrost thaw and subsidence of polygonal ground preceded and facilitated the increased inundation and sedimentation, or (3) these processes were both active.
Under scenario (1), and assuming relatively thin sediment deposits such as those observed here, the occurrence of larger or more frequent storm surges (Arp et al 2010) and onset of regular inundation at these locations would have killed existing vegetation and dampened the soil, leading to an increase in soil thermal conductivity, followed by permafrost thaw and ground subsidence (figures 4 and 5; Jorgenson et al 2010, O'Donnell et al 2009). Subsidence increases the area susceptible to subsequent flooding, thereby expanding the halophytic sedge habitat as we observed.
Scenario (2), where subsidence precedes flooding, requires that heat be transferred to the undisturbed permafrost from the adjacent degraded polygonal ground or from warmer summer temperatures or increased precipitation (Bhatt et al 2010, Jorgenson et al 2006). A study using interferometric SAR conducted 115-km east of our study area over a large swath of the Arctic Coastal Plain showed 1–4 cm of subsidence during the 1990s, which was attributed to thawing the top of the ice-rich permafrost (Liu et al 2010). If their reported average subsidence of 2 cm/decade also occurred at Smith River and Garry Creek, then it may explain our observed 4–7 cm sedimentation or vegetation aggradation since 1973. Using similar time series imagery as ours, ice wedge thaw was observed in inland polygonal terrain (Jorgenson et al 2006), and comparable degradation appears to have occurred at scattered locations throughout our study area (figure 5), but our sampling did not directly address permafrost thaw where marine flooding was absent.
Scenario (3) includes some combination of inundation (either salt or fresh water) and regional warming causing permafrost thaw and subsidence. Because our results only demonstrate the outcome, the relative importance of the different mechanisms remains unknown. Modeling suggests that processes in the marine environment are indeed acting in tandem with near-shore permafrost dynamics (Lawrence et al 2008). Increased marine inundation is likely related to climate warming through reduced sea ice extent, and related changes in storm frequency and intensity (Arp et al 2010). The same warming probably explains the regional subsidence observed to the east (Liu et al 2010). Thus, we believe the most likely explanation of our results is some combination of increased inundation with permafrost thaw and subsidence (figure 7). Regardless of the triggering mechanism, once regular inundation occurs, the thermal conductivity increases dramatically, which induces permafrost thaw, subsidence, and sedimentation.
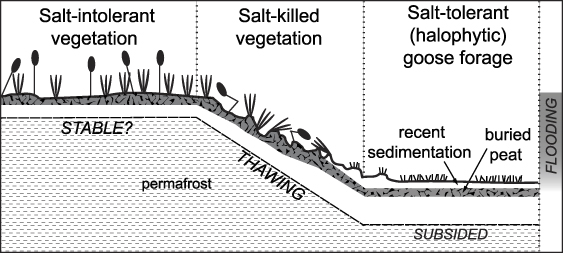
Figure 7. Idealized cross-section of the Garry Creek estuary on the Arctic Coastal Plain of Alaska. The diagram illustrates the proposed relationships between inundation, permafrost thaw, subsidence, and sedimentation, which lead to development of halophytic vegetation. Figure 4 contains a picture of this interface.
Download figure:
Standard image High-resolution imageThe process of inundation, subsidence, sedimentation, and goose habitat formation is ongoing and may be accelerating under conditions of sea ice decline, marine storm surge or freshwater flooding, and accelerated coastal erosion. The emigration of large numbers of Brant from inland habitats to coastal ones is consistent with our results showing that Brant habitat expanded near the coast. Recent survey data indicate that the higher numbers of Brant near the Smith River and Garry Creek are also occurring at other coastal locations (i.e., Piasuk River Delta and Cape Simpson; Flint et al 2013); these areas comprise more than 100 km of coastline. We suspect that our results are therefore not unique, and that geese in these low-relief coastal ecosystems are similarly responding to an increase in grazing habitat associated with new sedimentation.
The documented acceleration of coastal retreat (Jones et al 2009), the evidence presented here of inundation and subsidence of inland areas, and the resulting influx of Brant to halophytic coastal habitats (Flint et al 2008) may portend an approaching 'tipping point' (Scheffer et al 2001) in these coastal ecosystems. The threshold here is a literal one—elevation—that maintains tundra vegetation sufficiently above sea level, but the complexities associated with permafrost–climate–sea ice–vegetation dynamics leave considerable uncertainty regarding future land elevation. Eclipsing this threshold seemingly hinges on whether the regional subsidence documented east of our study area, apparently resulting from permafrost thaw (Liu et al 2010), is occurring in other portions of coastal and interior TLSA, and whether subsidence is outpaced by coastal erosion. Where subsidence and sedimentation outpaces coastal erosion, as suggested here, halophytic vegetation expands, and a threshold is crossed that initially expands foraging habitat and attracts molting and breeding geese of multiple species. A wider band of halophytic vegetation extending inland from the coast, such as documented herein, would constitute a conversion from a freshwater to marine-influenced ecosystem; once converted to sparse halophytic vegetation and regular inundation, the ecosystem would be unlikely to rebuild the intermediate layer of permafrost necessary to elevate the surface and revert it to a freshwater system. We suggest that future work examine elevation changes, with particular attention to low-lying areas near the coast and adjacent to floodplains that are susceptible to subsidence and inundation.
Acknowledgments
We thank Roger Ruess and John Pearce for suggestions that improved this letter. This work is part of the US Geological Survey's Changing Arctic Ecosystem Initiative and is supported by funding from the Wildlife Program of the USGS Ecosystem Mission Area. Use of any trade names in this letter does not imply endorsement by the US Government.
Appendix
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure A.1. Soil 14C values used for reconstructing bomb-curve chronology within soil pits at the Smith River estuary on the Arctic Coastal Plain of Alaska. Open symbols denote data that were ignored in fitting the 14C peaks to the bomb-curve because they were not post-modern or not part of an identifiable ascending or descending slope; nomenclature refers to soil profiles (figure 3).
Download figure:
Standard image High-resolution image{kind=link}