
Abstract
Corneal transplantation is an important surgical treatment for many common corneal diseases. However, a worldwide shortage of tissue from suitable corneal donors has meant that many people are not able to receive sight-restoring operations. In addition, rejection is a major cause of corneal transplant failure. Bioengineering corneal tissue has recently gained widespread attention. In order to facilitate corneal regeneration, a range of materials is currently being investigated. The ideal substrate requires sufficient tectonic durability, biocompatibility with cultured cellular elements, transparency, and perhaps biodegradability and clinical compliance. This review considers the anatomy and function of the native cornea as a precursor to evaluating a variety of biomaterials for corneal regeneration including key characteristics for optimal material form and function. The integration of appropriate cells with the most appropriate biomaterials is also discussed. Taken together, the information provided offers insight into the requirements for fabricating synthetic and semisynthetic corneas for in vitro modeling of tissue development and disease, pharmaceutical screening, and in vivo application for regenerative medicine.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
The cornea is a transparent, avascular and highly innervated tissue located at the anterior part of the eye. Its primary functions are to transmit and refract light entering the eye and to protect the eye from mechanical damage, UV light, and infection. Corneal transplantation is an important surgical treatment for injury and disease [1]. The World Health Organization (WHO) 2010 global survey estimated 39 million people with blindness. 12% or 4.9 million have bilateral corneal blindness [2]. From regional surveys, 23 million people are estimated to suffer from unilateral corneal blindness globally [3]. The causes of corneal blindness are diverse and include various corneal dystrophies, infectious and inflammatory corneal disorders, and corneal surface damage.
Corneal transplantation remains an important treatment for corneal diseases/conditions, with 72% of recipients showing visual improvement [4]. However, several unfavorable factors impede the utilization of corneal transplantation. In particular, although allogeneic tissue from human donors remains the best option for corneal graft replacement, this approach is hindered by the shortage of donor tissue and by transplant rejection [5].
A global survey on corneal transplantation showed there were a total of 184 576 corneal transplantations conducted in 116 countries and 284 000 corneas procured in 2012 [6]. It also reported a substantial lack of tissue with only one cornea available for every 70 patients worldwide [6]. The incidence of immunologic corneal graft rejection has decreased compared with solid organ transplantation by virtue of the cornea being avascular, an immunosuppressive ocular microenvironment, and the phenomenon of anterior chamber-associated immune deviation. Nevertheless, a recent study during a five year follow-up shows 23% of subjects experienced at least one rejection event after penetrating keratoplasty (PK) [7]. Normally, corneal immunologic rejection can be reversed with topical steroids, but severe rejection can lead to fast and permanent loss of endothelial cells and transplant failure. If no clinical improvement occurs, the only option is to surgically remove and replace the damaged cornea with a further corneal transplant [5, 8]. What is more, corneal rejection can result in permanent blindness [9]. Therefore, allogeneic corneal transplantation has limitations and a bioengineered cornea could potentially address the key issues of tissue availability and tissue rejection.
The first full-thickness PK was performed on 7 December 1905 by Dr Eduard Zirm [9]. The first case of lamellar keratoplasty (LK; removal of anterior layers of the cornea) was successfully conducted prior to this, but early outcomes were poor [10]. With the development of microsurgical instruments, and improved surgical techniques in the past ten years, selective lamellar keratoplasty (SLK; removal of diseased layers of corneas only), as the successor to LK, has significantly improved keratoplasty [1]. Specifically, SLK consists of anterior lamellar keratoplasty (ALK) and endothelial keratoplasty (EK). ALK replaces the epithelium and stroma, while retaining the uninfected healthy stroma and endothelium. Alternatively, a deep anterior lamellar keratoplasty (DALK) is performed if almost all the stroma has to be removed. DALK has been successfully used in treating keratoconus patients [11]. In contrast, EK replaces only Descemet's membrane and the endothelium, and is mainly used for corneal disease caused by endothelial disorders or failures. Compared to PK, ALK and DALK are less likely to cause loss of the host endothelial cells and thereby avoid endothelial rejection. EK can theoretically provide better visual outcomes due to improved astigmatism [1]. Recently, the frequency of lamellar procedures has been steadily increasing [12]. Nonetheless, all the above-mentioned techniques use donor corneas and therefore graft tissue availability and rejection remain challenging.
Since the discovery of limbal stem cells (LSCs), autologous or allogenic limbal tissue transplantation have been considered for reconstructing the injured ocular surface [13]. For patients with unilateral limbal stem cell deficiency (LSCD), a limbal autograft from the healthy contralateral eye is the best option for avoiding tissue rejection [14]. By contrast, the treatment for severe bilateral ocular surface disease is largely dependent on procuring allogeneic tissue from the limbal region of corneal tissue donors [13]. Benefits aside, it remains difficult to estimate the appropriate amount of limbal tissue from a donor eye for autologous transplantation. Inadequate volume may lead to the failure of eye surface-reconstruction and excess extraction tends to impair the healthy donor eye. Amniotic membrane (AM) has commonly been used as a natural substrate for limbal epithelial cell culture and delivery, but the potential for infection and the shortage of source tissue necessitate better alternatives [15]. Similarly, for reversible blindness caused by corneal endothelial cell failure, there is an urgent need for cell carriers to replace the failed endothelium with ex vivo expanded endothelium cells [16].
In this review, we provide an overview of the anatomy and function of the native cornea as the primer to a systematic summary of various biomaterials and methods for corneal engineering, including key characteristics for optimal material form and function, and suitable cells for material interfacing.
2. Human cornea anatomy and function
The human cornea is the first tissue through which light enters the eye and accounts for two thirds of the refractive power of the eye, directing the light rays towards the pupil [17]. Anatomically, the cornea is wider along the horizontal meridian compared to vertically. Population studies indicate the average horizontal diameter of the cornea in adults is around 11.80 mm [18, 19], and the vertical diameter is around 10.63 mm [20]. The radius of the corneal curvature is steepest at the center and flattens towards the periphery, with a mean central radius of curvature being approximately 7.7 ± 0.30 mm [21]. Until recently, the cornea was understood to comprise five layers, including anterior to posterior: epithelium, Bowman's layer, stroma, Descemet's membrane and endothelium (figure 1). In 2013, Dua et al reported a sixth layer termed pre-Descemet's layer that lies between the stroma and Descemet's membrane [22], although there is still controversy around its existence. Among these layers, the epithelium, stroma and endothelium are cellular layers, whereas Bowman's layer, pre-Descemet's layer and Descemet's membrane are acellular layers (figure 4).

Figure 1. Cross-sectional schema of the cornea.
Download figure:
Standard image High-resolution imageThe cornea is one of the most innervated tissues of the body, with the majority of nerves being sensory and derived from the ophthalmic division of the trigeminal nerve (5th cranial nerve) [23]. The nerve bundles enter the cornea through the limbus where they form the limbal plexus, then extend to the peripheral corneal stroma. The stromal nerve bundles start to branch at the mid-peripheral area, passing through Bowman's layer forming the sub-basal nerve plexus that locates in the basal epithelial layers. Individual nerve fibers arising from the sub-basal plexus move anteriorly and terminate in the superficial epithelium as free nerve endings [23]. Corneal nerves are important for both protection and maintaining the integrity of the ocular surface, and corneal sensation and nerve density are important indicators for cornea recovery after surgery [23, 24].
2.1. Corneal epithelium
The corneal epithelium is the outermost layer of the cornea and the major refractive element of the eye. It is 40–50 μm thick and consists of 4–6 layers of stratified and nonkeratinized squamous epithelial cells [17] (figure 2). The most superficial corneal epithelial cells form 2 ∼ 3 flat layers with glycocalyx-covered microvilli on the top surface. The glycocalyx is indispensable for the stability of tear film, which in turn contributes to normal vision. These surface cells form tight junctions that prevent tears, toxins and microbes from entering the eye. Immediately below the superficial cells is the middle wing cell layer, and the inner basal columnar layer. The basal cell layer is approximately 20 mm thick and the source of wing and superficial cells and, apart from LSCs and transient amplifying cells, are the only corneal cells capable of mitosis [25]. Furthermore, the basal cells are firmly connected to each other by lateral gap junctions and zonulae adherens and strongly attached to underlying basal lamina by hemidesmosomes [25].

Figure 2. Cross-sectional schema of the corneal epithelium.
Download figure:
Standard image High-resolution imageCorneal epithelial cells regenerate every 7–10 days through centripetal migration starting from the limbus, located on the corneal rim at the junction between the sclera and cornea (figure 3) [26]. The limbus contains corneal epithelial stem cells that are important for corneal epithelial homeostasis (figure 4(C)). The stem cells divide asymmetrically to produce transient amplifying cells, which migrate towards the central cornea to become basal central corneal epithelial cells. These cells then further differentiate and migrate towards the corneal surface to become the wing and superficial corneal epithelial cells (figure 3) [27]. In the normal resting state, p63 is totally absent in the corneal epithelium. When wound healing occurs, the αisoform of ∇Np63 (N-terminally truncated transcripts generated by the p63 gene) was reported to be significant to identify the stem cells that reside in the basal layer of the limbus. ∇Np63α, is an indicator for the regenerative proliferation of the corneal epithelium and can also be detected in the central cornea due to the migration of LSCs once activated [28]. The main function of the corneal epithelium is to form a barrier to prevent toxins and microbes from entering the epithelium, integrate with tears to provide a smooth layer for light refraction, and transfer water and soluble components into or out of the stroma [29]. The most posterior layer of the epithelium is a basement membrane (BM), where the basal epithelial cells attach. The BM is about 40–60 nm thick and consists of type IV collagen, laminin, perlacan (a heparin sulfate proteoglycan), and nidogen (a sulfated glycoprotein) [30]. Together, the BM is essential for the adhesion and polarity of the epithelial cells, and to modulate cellular signaling and trafficking between the epithelium and stroma [30].

Figure 3. Schema of the LSC niche.
Download figure:
Standard image High-resolution image
Figure 4. Light-microscopic image of the human cornea stained with hematoxylin and eosin. A human cornea is uniformly thick across the central region with increased thickness towards the limbus. B. Cross-sectional image showing the six-layered corneal structure. C. Enlarged image of the limbus showing the presence of blood vessels and an undulating basal epithelium (asterisks).
Download figure:
Standard image High-resolution image2.2. Bowman's layer
Between the epithelium and stroma layers lies the Bowman's layer (figure 1), which has a smooth anterior surface abutting the epithelial BM, with its posterior side merging with the stroma [31]. The Bowman's layer is acellular and has numerous pores allowing the passage of nerve bundles [31]. It consists mainly of collagen types I and III, as well as types V and VI [32–34], which form collagen fibrils with a diameter of 20–25 μm. The individual collagen fibrils interconnect to generate a sheet about 8–12 μm [35]. The thickness of the Bowman's layer decreases with age [35] and it does not regenerate following trauma or removal [31]. It is hypothesized that the Bowman's layer is formed early in the development of the cornea via the interaction between the corneal epithelial cells and keratocytes in the stroma [36]. Its absence does not appear to prevent re-epithelialization following surgery or trauma [36]. However, the Bowman's layer has been reported to assist with stromal wound healing and restoration of the subepithelial nerve plexus [31].
2.3. Stroma
The corneal stroma accounts for roughly 80%–85% of the entire corneal thickness, and affords many biochemical properties of the cornea including tensile strength, stability and transparency [30]. The collagen fibrils in the stroma are bigger than in the Bowman's layer with diameters ranging from 25–35 nm [35]. Collagen type I is predominant in the stroma. Together with type III and V collagen they form collagen fibrils. Type VI collagen occurs as fine filaments in the inter-fibrillary matrix of the stroma [34]. The collagen fibrils form triple-helix bundles with diameters of 250–340 nm, which align to form a flat lamellar sheet [35]. The lamella sheets are enveloped by proteoglycans decorated with keratin sulfate or chondroitin sulfate, and preserve corneal hydration and transparency [37]. The stroma consists of 250 ∼ 300 collagen lamellae with a width of 10–320 μm and a thickness of 0.2–2.5 μm [38]. The arrangement of the lamellae in the stroma is heterogeneous, with the collagen fibers being interwoven in the anterior stroma and parallel in the mid to posterior stroma [38]. In addition, the intervals between lamellae are filled with keratocytes, which occupy more than 20% of the stroma. These cells are responsible for the stability of lamellar organization and regulation of the corneal stroma extracellular matrix (ECM) components via the synthesis of collagen and proteoglycan [39]. The framework of the corneal stroma is important for the transparency and mechanical property of the cornea.
Once the normal integrity or the interconnection with the Bowman's layer and Descemet's membrane is interrupted, the stromal wound-healing process is activated. Due to the distinct avascular nature of the cornea, a critical difference between stromal healing and that of other body tissues such as the skin is the absence of the vascular component [40].
2.4. Pre-Descemet's layer and Descemet's membrane
The pre-Descemet's layer is posterior to the stroma and anterior to the Descemet's membrane. The recent discovery of the pre-Descemet's layer remains controversial with many studies suggesting the layer to be part of the stroma [22, 41]. However, emerging evidence of the structure, constitution and clinical importance support its existence as a distinct layer. It is made up predominately of type I collagen but includes type VI collagen, which assembles into 5–8 thin lamellae with an overall thickness of 10–15 μm [22]. A recent study suggests that it consists of high levels of elastic fibers that originate within the posterior limbus in the trabecular meshwork [42]. This elastic fiber network in the pre-Descemet's layer could be important for anchoring the cornea as well as the trabecular meshwork to the limbus and peripheral cornea [42]. The clinical importance of the pre-Descemet's layer is supported by the big bubble technique being unsuccessful if the layer is disrupted [43]. The predominant collagen in the Descemet's membrane is type IV, although it also includes collagen type VIII, XII, laminin, perlecan, nidogens, vitronection and fibronectin [44]. In contrast to the Bowman's layer, its thickness gradually increases from birth to adulthood, expanding to between 5–10 μm [45]. It is secreted by the endothelial cells to form the BM for the endothelial cells to anchor [45].
2.5. Endothelium
The human corneal endothelium (HCEN) is composed of a single layer of hexagonal-shaped cells with a thickness of around 5 μm. Similarly, the adjacent endothelial cells are characterized by a lateral gap and tight junctions and are vertically attached to the Descemet's membrane by numerous hemidesmosomes [46]. Once the endothelium layer is formed, the endothelial cells become mitotically inactive. The HCEN initially comprises approximately 4500 cells mm−2, with the total endothelial cell number decreasing with age, while the cell surface area increases [47]. The main indicator for an eye bank to select corneas after storage for PK, is the endothelial cell counts [48]. Generally, donor corneas considered for transplantation need to have an endothelial cell density of ≥2000 cells mm−2.
While acting as a barrier to restrain fluid into the cornea, the endothelial cells prevent excessive hydration (78% water content) of the stroma, maintaining stromal deturgescence by a passive pump-leak process that moves ions and water from the hypotonic stroma to the hypertonic aqueous humor [49]. The whole process has been shown to be associated with Na+, K+– adenosine triphosphatase (ATPase) pumps that reside in the basolateral membrane [50].
Thus, the endothelium is critical to maintaining the transparency and nutrition of the cornea via this 'pump-leak' mechanism. It is rich in ion and water channels for the transfer of nutrients from the aqueous humor to the cornea [51] and its function to nourish the cornea and inability to regenerate, renders it key to determining by assessment the suitability of the cornea for transplantation.
Due to their mitotically inactive nature, once the number of endothelial cells decreases, the surrounding cells gradually infiltrate the vacant region to maintain tissue integrity. The deformation of the endothelium, however, tends to impair the pump function [52].
3. Biomaterials for corneal engineering
Biomaterials employed for engineering corneal substitutes must replicate the structural and functional requirements of the native cornea. Briefly, as scaffolds for cell support, materials must have suitable mechanical toughness, biocompatibility, transparency, appropriate biodegradability, and be clinically compliant. Commonly used biomaterials broadly include natural and synthetic polymers. While natural polymers tend to have excellent biocompatibility, synthetic polymers enable customization of the desired properties. The characteristics of extant materials determine their application (tables 1 and 2).
Table 1. Biomaterials used for corneal bioengineering.
| Material | N/Sl | Processing method | Pros | Cons | Applications | Biocompatibility | Transparency | Mechanical toughness | Biodegradability | Permeability | Clinical status |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Collagen | N | Crosslinking; | High biocompatibility from intrinsic RGDm | Inferior mechanical properties | Corneal stromal equivalents; | ++++ | +++ | + | ++++ | +++ | Clinical follow-up 48 months |
| Collagen-vitrigel; | Substrate for corneal epithelial and endothelial bioengineering | ||||||||||
| Compression | |||||||||||
| Silk | N | Evaporation; | Inherent optical clarity; controllable degradation rates and mechanical properties | Surface modification or combination required | Corneal stromal equivalents; substrate for corneal epithelial cells; corneal fibroblasts and corneal endothelial cells | ++ | ++++ | +++ | +++ | +++ | Animal model |
| Electrospinning | |||||||||||
| Gelatin | N | Evaporation; | Natural biocompatibility; suitable biodegradability | Inferior | Substrate for | +++ | +++ | + | ++++ | +++ | Animal |
| Crosslinking | mechanical properties | corneal endothelial cell transplantation and corneal stromal cell growth | model | ||||||||
| Chitosan | N | Evaporation; | Easily-biofunctionalized; favorable biocompatibility; controllable biodegradability | Composition of other materials (collagen /gelatin) required | Substrate for corneal epithelial regeneration and delivery and for endothelial cell growth | ++ | +++ | ++ | ++++ | ++++ | Animal model |
| Composition and crosslinking | |||||||||||
| DCa | N | Decellularization | Similar mechanical and optical properties to natural cornea; low immunogenicity | Insufficient keratocyte infiltration and bioactivity | Substrate for corneal endothelial regeneration and epithelial and anterior stromal reconstruction | ++ | ++++ | +++ | +++ | None | Animal model |
| SSb | N | Coating; evaporation | High biocompatibility | Inferior mechanical property | Substrate for human corneal limbal epithelial cell | +++ | None | ++ | None | None | In vitro |
| Fibrin/Agarose | N | Polymerization | Excellent optical properties | Complicated process | Keratocytes encapsulated corneal stroma equivalent | +++ | ++++ | None | None | None | In vitro |
| PVAc | S | Crosslinking; Electrospinning | Sufficient mechanical properties | Surface modification required; unsatisfied light transmittance | Substrate for corneal epithelial and stromal regeneration and delivery | + | + | None | ++ | None | In vitro |
| PHEMAd | S | Crosslinking | Favorable transparency | Surface modification required | Substrate for limbal epithelial cells | + | ++++ | ++++ | + | None | In vitro |
| PEGDAe | S | Crosslinking | Controlled biochemical and topographical cues; low relaxed swelling ratios | Surface modification required | Substrate for corneal epithelial cells; epithelial wound healing | None | None | None | None | None | In vitro |
| PLGAf | S | Evaporation | High transparency | Surface modification required | Substrate for corneal endothelial regeneration | + | ++++ | None | +++ | None | In vitro |
| PEGg/PAAh hydrogel | S | Polymerization and crosslinking | High transparency and glucose permeability | Complicated process | Corneal stroma and epithelial reconstruction | ++ | ++++ | ++ | ++ | ++++ | Animal model |
| PGSi | S | Polycondensation | Favorable biodegradability | Complicated process | Substrate for human corneal epithelial cells | ++ | None | None | None | +++ | None |
| PEUUj | S | Electrospinning | Excellent biodegradability | Complicated process | Substrate for corneal stromal cell growth and regeneration | ++ | None | None | ++++ | None | In vitro |
| Thermoresponsive materials | S | Graft; coating | Generate carrier-free cell sheet without enzymatic digestion | Generated cell layers are too fragile for surgical manipulation | Substrate for corneal endothelial sheets and epithelial sheets | ++ | None | None | None | None | Animal model (Cell sheets) |
| PAk | S | Self-assembly (in solution and coating) | Versatile and easily designed | Complicated process | Design 2D and 3D structures for corneal cells and improve cell performance | ++++ | None | None | +++ | None | Animal model |
| biocompatibility and structure |
aDecellurized cornea. bSilk sericin. cPoly(vinyl alcohol). dPoly(2-hydroxyethyl methacrylate). ePolyethylene (glycol) diacrylate. fPoly(lactic-co-glycolic acid). gPoly(ethylene glycol). hPoly(acrylic acid). iPoly(glycerol sebacate). jPoly(ester urethane) urea. kPeptide amphiphiles. lNaturally/synthetically derived. mArginine-glycine-aspartic acid. '++++' stands for the highest level, while '+' is the lowest.
Table 2. Biomaterials and cell sources used for corneal bioengineering for each cellular layer.
| Materials | Cell sources | |
|---|---|---|
| Epithelium | Silk fibroin: | Primary human corneal limbal epithelial cells |
| Silk fibroin membrane [93, 95, 100] | [56, 70, 72, 79, 87, 89, 90, 93, 95, 100, 104, 107, 121, 129, 141, 152, 160] | |
| Porous silk fibroin membrane [104, 190] | ||
| Patterned silk fibroin membrane [152] | ||
| Silk sericin [107] | ||
| Collagen: | Primary human corneal epithelial cells | |
| Type I collagen gel [55, 59, 83, 89] | [78, 80, 89, 142, 143, 151, 154, 190] | |
| FNCa [54] | Immortalized human corneal epithelial cells | |
| CVb [56, 86] | [54, 59, 60, 67, 71, 82, 83, 129, 148, 150] | |
| Collagen/silk gel [82] | ||
| Synthetic hydrogel | Immortalized human corneal limbal epithelial cells [153] | |
| PHEMAc [141] | ||
| PEGDAd [142, 143, 159] | ||
| Decellurized cornea [121, 125] | ||
| Chitosan [128] | Primary rabbit corneal limbal epithelial cells [55, 117, 163, 166, 194] | |
| Hydroxyethyl chitosan [133] | ||
| Thermoresponsive polymer PNIPAAme [134] | Primary rabbit cornea epithelial cells [133, 140, 165] | |
| Electrospun scaffold: | Primary bovine corneal epithelial cells [128] | |
| PLGAf [163] PHBVg/gelatin [111] | Primary porcine corneal limbal epithelial cells [152] | |
| Chitosan/gollagen [129] PCLh/PVAi [166] | ||
| Gelatin/PLLAj [165] | ||
| Stroma | Silk fibroin: | Primary human corneal stromal stem cells [164, 170, 181] |
| Porous silk fibroin membrane [105, 190] | ||
| Patterned silk fibroin membrane [96, 97, 101, 158] | Primary human corneal fibroblasts [58, 62, 89, 121, 152, 173–175, 181] | |
| RGDk functionalized silk film [97, 105] | ||
| Collagen: | ||
| Type I collagen [65, 89, 92] | Primary human corneal keratocytes [59, 79, 90–92, 155, 158] | |
| Insoluble type I collagen [65] | ||
| CV [56] | ||
| PEUUl [164, 180, 181] | Immortalized human corneal keratocytes [71, 96, 97, 101, 176] | |
| Gelatin hydrogel [111, 113] | ||
| DCm [121, 122] | ||
| PLDLAn [155] | Primary rabbit corneal stromal cells | |
| Electrospun scaffold: | [106, 113, 117, 122, 140, 162, 165, 194] | |
| PHBo/PHBV/PCL [162] | ||
| PEUU [164] | Primary rabbit corneal fibroblasts precursors [111] | |
| Gelatin/PLLA [165] | Primary rabbit corneal stromal fibroblasts [111] | |
| Other animal corneal stromal cells: goat [157]; bovine [148] | ||
| Endothelium | Gelatin hydrogel [110, 112, 114] | Primary human corneal endothelial cells |
| Gelma [116] | [64, 90, 98, 110, 114, 116, 123, 136, 170] | |
| Thermoresponsive polymer: | ||
| PNIPAAm [136] | Immobilized human corneal endothelial cells [71, 79, 98, 138, 139] | |
| Poly (NiPAAm-co-DEGMA)p [138] | ||
| PNIPAAm hybrids [157] | ||
| Poly(NGMA)q [137] | Primary rabbit corneal endothelial cells [56, 102, 112, 117, 131, 132, 137, 144, 194] | |
| PVMEr [139] | ||
| Chitosan [131] | ||
| HECTSs [132] | Primary bovine corneal endothelial cells [130, 156] | |
| PCL-chitosan [130] | ||
| Silk fibroin [98] | ||
| Aloe vera gel/SF [102] | ||
| DC [123] | ||
| Compressed collagen [64] | ||
| Full-thickness | Silk fibroin [38] | Dorsal root ganglion (DRG) cells: |
| Fibrin [192, 193] | from chick embryos [66, 76, 190] | |
| Collagen gel: | ||
| Type I collagen [60] | Human embryonic stem cells (hESCs) [127] | |
| Type I and type III collagen [66] | ||
| Collagen I and copolymer [74–77] [79] | ||
| CV [57] | ||
| DC [117, 118, 126, 127] | ||
| PEGt/PAAu hydrogel [146, 147] | ||
| PHEMA/PAA hydrogel [145] | ||
aFibronectin-collagen. bCollagen-vitrigel. cPoly(2-hydroxyethyl methacrylate). dPoly(ethylene glycol) diacrylate. ePoly(N-isopropylacrylamide). fPoly(lactide-co-glycolide). gPoly(3-hydroxybutyrate-co-3-hydroxyvalerate). hPoly(ε-caprolactone). iPolyvinyl alcohol. jPoly-L-lactic acid. kArg-Gly-Asp. lPoly(ester urethane) urea. mDecellurized cornea. nPoly(L,D lactic acid). oPolyhydroxybutyrate. pPoly(N-isopropylacrylamide-co-diethyleneglycol methacrylate. qPoly(N-isopropylacrylamide-co-glycidylmethacrylate). rPoly(vinyl methyl ether). sHydroxyethyl chitosan. tPolyethylene glycol. uPoly(acrylic acid).
3.1. Collagen
Collagen is the most abundant component of ECM in most tissues [53, 54], with tripeptide arginine-glycine-aspartic acid (RGD) recognized by the cell surface integrin receptors, being important for cell adhesion, migration and proliferation [55]. The widespread application of collagen (especially collagen type I) in tissue engineering is partly due to its ease of production. Collagen type I is a key component to the tendon, ligament and dermis and is relatively cheap to derive. Collagen is therefore vital to fabricating biomimetic corneal stroma equivalents as the main component of human stromal tissue [56–58], and is compatible with all corneal cells and nerve regrowth in vivo [55, 59–62]. The primary drawback of collagen, however, is insufficient mechanical toughness and elasticity [63, 64]. Therefore, crosslinking collagen has been widely applied to considerably improve its mechanical strength and chemical stability without compromising the biological advantages. Importantly, the source of collagen impacts on the physical properties of the final product. For instance, the addition of soluble tropocollagen improves transparency and strength [65], and while both type I and type III collagen hydrogels have adequate tensile strength and elasticity for handling, type III collagen hydrogels tend to be mechanically and optically superior [66, 67].
Table 3. Collagen used for corneal bioengineering.
| Collagen Type | Processing method | References | Year | In vitro study | In vivo study |
|---|---|---|---|---|---|
| Bovine type I collagen | Dehydrothermal crosslinking | [89] | 1999 | Primary human corneal epithelial cells (limbus and center) and human corneal fibroblasts | None |
| Bovine type I collagen | Dehydrothermal crosslinking | [58] | 2004 | Primary human corneal stromal fibroblasts | None |
| Bovine collagen type I | Dehydrothermal crosslinking | [62] | 2006 | Primary human corneal stromal fibroblasts | None |
| Bovine type I collagen and type III collagen | Glutaraldehyde crosslinking; EDCa/NHSii crosslinking | [59] | 2006 | Immortalized human corneal epithelial cells | None |
| Porcine type I collagen | EDC/NHS crosslinking | [61] | 2006 | None | Rabbits and mini-pigs by lamellar keratoplasty |
| Bovine collagen type I | EDC/NHS crosslinking | [83] | 2007 | Immortalized human corneal epithelial cells | None |
| Recombinant human collagens types I and III | EDC/NHS crosslinking | [66] | 2008 | Immortalized human corneal epithelial cells | Mini-pigs by deep lamellar keratoplasty |
| Recombinant human collagens types I and III | EDC/NHS crosslinking | [67] | 2008 | Immortalized human corneal epithelial cells | Rats by selective lamellar keratoplasty |
| Rat tail type I collagen | EDC/NHS crosslinking | [79] | 2008 | Primary cornea limbal epithelial cells; primary human keratocytes; immortalized human endothelial cells; | None |
| Type I porcine atelocollagen; recombinant human type III collagen | EDC/NHS crosslinking | [76] | 2009 | Immortalized human corneal epithelial cell line and dorsal root ganglia | Rabbits by deep lamellar keratoplasty |
| Type I collagen solution | Collagen-vitrigel | [56] | 2009 | Primary human limbal epithelial cells; primary bovine keratocytes; rabbit corneal endothelial cells | None |
| Recombinant human collagen, type III | EDC/NHS crosslinking | [177] | 2010 | None | 24 month follow-up of a phase 1 clinical study |
| Type I porcine atelocollagen | EDC/NHS crosslinking | [77] | 2010 | Immortalized human epithelial cell line and dorsal root ganglia | Pigs by penetrating keratoplasty |
| Rat tail type I collagen | Dehydrothermal crosslinking | [87] | 2012 | Primary human limbal epithelial stem cells and stromal keratocytes | Rabbits by selective lamellar keratoplasty |
| Rat tail type I collagen solution | Dehydrothermal crosslinking | [55] | 2011 | Rabbit limbal epithelial cell | None |
| Rat tail type I collagen | Dehydrothermal crosslinking and plastic compression | [81] | 2014 | Primary human corneal endothelial cells; human corneal endothelial cell line | None |
| Fibronectin-collagen treatment | [54] | 2013 | Immortalized human corneal epithelial cells | None | |
| Bovine type I collagen | EDC/NHS crosslinking | [154] | 2014 | Primary human corneal epithelial cells | None |
| Bovine type I collagen | EDC/NHS crosslinking | [78] | 2014 | Primary human corneal epithelial cells | None |
| Bovine type I collagen | EDC/NHS crosslinking | [82] | 2015 | Human corneal epithelial cells | Rabbits by lamellar keratoplasty |
| Bovine type I atelocollagen | Collagen-vitrigel | [86] | 2015 | None | Rabbits by lamellar keratoplasty and LSCDiii model induced by chemical injuries. |
| Rat collagen type I | Dehydrothermal crosslinking | [70] | 2015 | None | Pig keratocyte; primary human limbal epithelial cells |
-Ethyl-3-(3-dimethylaminopropyl)-carbodiimide. ii N-hydroxysulfosuccinimide. iiiLSC deficiency.
The methods which have been investigated for collagen crosslinking for bioengineering mainly include chemical, physical and enzymatic approaches. Aldehydes such as formaldehyde and glutaraldehyde, the carbodiimide family (EDC), isocyanate chemical family (hexamethylene diisocyanate) and genipin are commonly used for the chemical crosslinking of collagen [68]. Physical crosslinking involves ultraviolet (UV) or dehydrothermal treatment and enzymatic crosslinking with transglutaminase can enhance the mechanical toughness of collagen without generating toxic by-products [68]. However, not all of these methods are commonly employed in corneal bioengineering, with some approaches generating toxic residues in the collagen-based biomaterial. Unlike glutaraldehyde, hexamethylene diisocyanate and acyl azide, which act as chemical 'bridges' between collagen molecules, with EDC, collagen molecules are linked directly. Genipin is a crosslinker with low toxicity [69]. A study of various crosslinking methods demonstrated the inclusion of generation 2 polypropyleneimine octamine dendrimers largely amplifies the quantity of amine groups in collagens available for EDC crosslinking. The membrane produced had superior optical and mechanical properties compared with dehydrothermally and glutaraldehyde crosslinked collagen gels [59]. Another study of collagen suggested that while UV light induced riboflavin, crosslinking or the use of genipin, or EDC crosslinking produced adequate tensile strength, superior strength was found using the EDC crosslinking group [70]. Another crosslinking method involving UV-induced glucose, results in collagen membranes with moduli and ultimate tensile strength comparable to that of the native human cornea [65].
Normally, crosslinked collagen gel can be produced in a membrane form that is suitable as a growth substrate for corneal cells [60] and biomimetic corneal equivalents [56, 57], or 3D structures with stromal cells encapsulated or scaffolds seeded with epithelial and endothelial cells [71, 72]. The latter provides for the development of full-thickness corneal tissue that is analogous to a native structure, but has relatively poor mechanical properties due to incomplete crosslinking and cell protection [71, 72]. The cytocompatibility of crosslinked collagen gels has been demonstrated in numerous studies. For example, dehydrothermally crosslinked, EDC and glutaraldehyde crosslinked gels are able to support the growth of human and rabbit corneal epithelial cells (rCECs), HCFs, and HCEN cells [55, 58, 60, 62, 71, 73]. In addition to pure collagen hydrogel, collagen-polymer composites have been reported to further enhance the mechanical strength and resistance to enzymatic degradation [74, 75]. For example, porcine type I collagen and 2-methacryloyloxyethyl phosphorylcholine (MPC) interpenetrating network hydrogels display improved mechanical strength, and stability, with comparable optical properties, and glucose as well as albumin permeability with those of the native human cornea [76, 77] enabling the regeneration of corneal cells and functional nerves in pig models. A combination of collagen with a copolymer from N-isopropylacrylamide, acrylic acid and acryloxysuccinimide resulted in a transparent, permeable and robust collagen gel that prevented excessive collagen fibrillogenesis and was well tolerated in dogs [74]. A composite collagen-chitosan membrane demonstrated better mechanical toughness and optical properties and when transplanted into a pig cornea, allowed the regeneration of the epithelium, stroma and nerve in 12 months [63, 78]. The incorporation of chondroitin sulfate into collagen gels tends to lead to higher mechanical properties and moisture retention [73, 79]. Crosslinking tobramycin into collagen, a novel antibacterial collagen membrane with excellent antibacterial effect can be obtained [80]. The incorporation of cyclodextrin in collagen membrane is capable of improving the transparency as a result of reduced collagen fibrogenesis [81]. Enhanced mechanical properties can also be achieved by the integration of silk fibroin [82]. Duan et al reported that the incorporation of YIGSR (a model cell adhesion peptide) modified dendrimer into collagen gel promoted the adhesion and proliferation of human corneal epithelial cells (HCECs) as well as neurite extension from dorsal root ganglia [83]. While most collagen structures reported are flat, Zhang et al recently produced a natural corneal-shaped collagen membrane by placing dethermally crosslinked collagen in a rigid contact lens mold and then incubated it at 37 °C for complete dehydration [70]. By loading a compressive force on the dehydrothermally crosslinked collagen gel, plastic compression (PC) collagen with improved mechanical toughness has been explored as a carrier to expand the HCEN cell for transplantation [84]. The PC collagen films, however, are not easily manipulated. To solve this issue, electrospun PLGA films were then used to form a sandwich-like structure for transplantation. The light transmittance of the structure, however, is insufficient, being less than 80% under a 500 nm wavelength [85]. Other composites include collagen-vitrigel (CV), a collagen-based product with enhanced gel strength by vitrification [86]. Briefly, the mixture of collagen, FBS, N-2-hydroxyethylpiperadine-N0-2-ethansulfonic acid buffer and Dulbecco's modified eagle medium was incubated at 37 °C and 5% CO2 for 2 h gelation. Under controlled humidity, the gel was vitrified into a glass-like material. Finally, the CVs were rehydrated to obtain a regenerated and stable CV membrane. During the whole process, the CV can be tailored for transparency and mechanical strength by optimizing the synthesis conditions [87]. Moreover, the CV has been shown to facilitate epithelial layer generation in vitro [56], and exhibits sustained tissue transparency and low inflammatory response [86].
To date, collagen is the most extensively used material in corneal bioengineering (table 3). Overall, collagen scaffolds, either pure or hybrid, provide excellent support of cells derived from different corneal tissue layers. The main challenge for tissue engineers using collagen for corneal bioengineering is the fabrication of collagen constructs that mimic the well-organized structure and cell-encapsulation ability of the corneal stroma [88]. Griffith et al created a multilayered corneal matrix by seeding epithelial cells and endothelial cells on the top and bottom of immobilized human corneal keratocyte-encapsulated glutaraldehyde crosslinked collagen-chondroitin sulfate substrate [71]. Similarly, German et al reconstructed a cornea by seeding primary HCECs on primary human fibroblasts encapsulated dehydrothermally into a crosslinked collagen gel. After 3 days in culture, 4–5 layers of the regenerated corneal epithelium and components of BM were detected [89]. Keratocytes have been incorporated into porous collagen sponges by crosslinking and lyophilisation [90]. The keratocytes migrated into the pores and evidence of matrix secretion and elongated cell morphology was observed. Controlling the organization of collagen fibrils to mimic the corneal stroma has long been of interest when utilizing collagen for corneal bioengineering. Builles et al reported that scaffolds of orthogonal lamellae composed of aligned collagen fibrils can be formed by the gelation of collagen in a horizontal magnetic field, mirroring the inner structure of the human corneal stroma (HCS) [91, 92]. Interestingly, keratocyte alignment is directed by the collagen orientation both on the surface and within the bulk of the scaffold [92]. Although the scaffold possesses similar structure to human cornea stroma, it is not transparent probably due to the relatively large diameter of collagen fibrils. Proteoglycans were thus incorporated into the scaffold to improve the transparency. Although the as-prepared scaffolds demonstrated great potential for the recovery of the anterior cornea in the rabbit model, the graft retention, however, still needed to be improved through the optimization of crosslinking [92].
3.2. Silk
Silk fibroin (SF) is a structural protein derived from the cocoon of the silkworm bombyx mori and has been widely used in tissue engineering and regenerative medicine, because of its non-immunogenic response, controllable degradation rates and mechanical properties. Normally, pure silk solution, the precursor of the diverse SF framework, is obtained by mixing SF with lithium bromide solution before dialysis. In the context of corneal engineering, the inherent optical clarity of SF undoubtedly makes it a promising candidate. SF membranes have largely been investigated as substrates for corneal epithelial cells. Pure and permeable SF membrane generated by simple evaporation [93] or electrospinning [94] is able to support the formation of a confluent multilayered epithelium and the growth of human corneal limbal epithelial (HCLE) cells similar to the AM, which is the current standard substrate used for corneal epithelial cell transplantation. Although pure SF membrane provides for similar cell attachment of HCLE cells to tissue culture plastics, it is inferior to AM due to the lack of natural ECM proteins [95]. Nevertheless, the RGD sequence present in collagen can be coupled on the surface of the SF membrane to enhance the attachment and proliferation of HCLE cells, human corneal fibroblasts (HCFs) [96, 97], and human corneal endothelial (HCEN) cells [98] by mediating cell-substrate adhesion interactions. Interestingly, poly-D-lysine (PDL) provides comparable support to modified RGD [99]. For corneal stromal bioengineering, porous silk film has been shown to support the construction of a two-layer structure using HCLE cells and stromal cells [100], and human corneal stromal stem cells (hCSSCs) while secreted ECM adhered to the material [101]. Moreover, enhanced cell viability and well-maintained morphology of rabbit corneal endothelial (rCEN) cells have been reported with the incorporation of natural aloe vera (AV) gel with SF [102]. To improve molecular permeability and biodegradability, porous SF structures were prepared using the porosity inducing agent poly(ethylene glycol) (PEG). The porous structure induced by PEG, however, tends to reduce the growth and stratification of human limbal epithelial cell (HLEC) growth. It is also noteworthy that by having high porosity the membranes tend to be more fragile [100, 103, 104]. Nonetheless, implanted RGD-functionalized SF films have been shown to keep their integrity and transparency for half a year without immunogenic and neovascular responses in rabbit stroma [105] and notably a faster degradation rate can be achieved by introducing porosity [101] or combining propionamide [106].
While fibroin has been extensively investigated in corneal engineering, another protein from the domesticated silkworm silk sericin (SS) has been consistently overlooked due to suspected allergenic activity. Recent interest, however, in the biocompatibility of SS has resulted in its consideration as a cell growth substrate. Pure or blended SS membrane showed inferior mechanical properties, but enhanced HCLE attachment compared with SF, suggesting a potential application for corneal epithelium regeneration [107]. Recently, Applegate et al innovatively published a silk photocrosslinking method using flavin-mononucleotide (FMN), which is a water-soluble variant of riboflavin, to transfer silk solution to a highly elastic and transparent hydrogel. The materials involved in the process are all biologically and environmentally friendly, which makes this technique very promising for corneal bioengineering [108].
Overall, compared with collagen, the advantage of using silk is mainly due to the simplicity of production and modification, as well as the ease of patterning and preparing the porous structure, which profoundly impacts the cell behavior of all corneal cells. The depth and width of grooves, for instance, were found to affect HCF alignment. Moreover, building a multilayered corneal structure using stacked patterned porous silk films or sponge provides a useful 3D model, in spite of the technique requiring further improvement with the native cornea consisting of approximately 200 lamellae with a total thickness of 500 microns, while the stacked silk films have far fewer layers [109]. While chemical surface modification has been investigated to improve the biocompatibility, further research is necessary since the proteins in the natural cornea are varied and multifunctional. Most of the studies to date have focused on the correlation between surface pattern and porous structure, with further studies of in vivo performance (including the influence of porosity and patterning on biocompatibility) necessary.
3.3. Gelatin
Gelatin is a ubiquitous natural material derived through the hydrolysis of collagen. The application of gelatin for corneal engineering mainly involves constructing membranes for corneal cells through crosslinking. The applied methods to prepare gelatin hydrogel generally include dehydrothermal and chemical crosslinking. Being dehydrothermally crosslinked, gelatin is able to provide sheets with better transparency, elastic modulus, and albumin permeability compared with collagen. In addition, primary HCEN cells seeded on as-prepared gelatin showed normal expression levels of ZO-1, Na +/K + -ATPase, and N-cadherin, and achieved a continuous endothelial monolayer [110]. Mimura et al reported chemically crosslinked gelatin using glutaraldehyde, which was able to support rabbit fibroblast, fibroblast precursor adherence and ECM deposition, and was suitable for implantation into a rabbit stromal pocket [111]. Using EDC/NHS crosslinking and freeze-drying, gelatin membranes with varying porosity, Young's modulus and swelling ratio can be prepared by varying the concentration of gelatin and crosslinking parameters [112]. Compared with pure gelatin, the incorporation of chondroitin sulfate (CS) has been shown to promote the growth of primary rabbit corneal keratocytes (RCKs). The hybrid gelatin-CS gel, however, had lower Young's modulus and resistance against protease digestion [113]. By adding heparin during EDC crosslinking, Niu et al prepared an innovative transparent gelatin scaffold that has the capacity to enhance the basic fibroblast growth factor absorption and release kinetics [114]. Furthermore, the scaffold is sufficiently flexible to be folded for easier transplantation and HCEN cells can be grown on the scaffold, while maintaining their endothelial morphology and critical pumping function. Following transplantation in rabbits, the implanted scaffolds gradually fuse with the surrounding stroma [114]. Lai reported a 3D culture system for RCKs using hyaluronic acid (HA) functionlized gelatin microspheres that supported the large-scale growth of RCKs. Investigation of the biocompatibility of microcarriers indicated compatibility with rabbit corneal epithelial, stromal, and endothelial cells, and they were well tolerated in the anterior chamber of the rabbit eye [115]. Semisynthetic gelatin methacrylate (GelMA) has been increasingly used in tissue engineering due to its biocompatibility and more importantly, tunable physical characteristics produced through radical polymerization in the presence of a photoinitiator. Rizwan et al developed a hybrid crosslinked GelMA hydrogel (GelMA+) by incubating a prepolymer solution at 4 °C for 1 h before UV crosslinking (figure 5). The GelMA+ showed superior mechanical properties due to the smaller and more uniform distribution of pores and excellent support of HCEN cells. GelMA+ films seeded with HCECs and transplanted into a rabbit model were associated with high cell viability following transplantation [116].

Figure 5. Schematic diagram of the fabrication of regular gelatin methacrylate (GelMA) and GelMA with hybrid crosslinking (GelMA+) (Reprinted from [116], Copyright (2017), with permission from Elsevier).
Download figure:
Standard image High-resolution image3.4. Decellularized cornea
Decellularized cornea (DC) has shown potential for corneal scaffold fabrication due to the similar mechanical and optical properties when compared to a natural cornea [117, 118] and low immunogenicity [119, 120]. The shortage of suitable corneal tissue as a requirement for a human DC remains a limiting factor and therefore most studies have employed porcine corneas or bovine corneas as the source material. Previous studies have revealed the feasibility of using a DC to support corneal cell growth in vitro and to facilitate tissue regeneration in animal models [121–123]. Conventional approaches to decellularize cornea tissue involve the removal of the cellular component using non-ionic detergent (e.g. Triton X-100, Sigma, St. Louis, MO, USA), ionic detergents (e.g. sodium dodecyl sulfate (SDS) and SD), zwitterionic detergent (e.g. CHAPS), freezing-thawing, and osmotic shock with hypotonic or hypertonic solutions [118, 123, 124]. Comparison between Triton X-100, SDS, SD and CHAPS revealed SDS to be the most effective for the removal of cellular components. Furthermore, 0.5% SDS was the most optimal concentration for cell removal, while preserving collagen ECM [118, 124]. Another study of decellularization using Triton X-100, liquid nitrogen, and poly(ethylene glycol), sodium dodecyl sulfate (SDS) and sodium chloride (NaCl) with nucleases found that only NaCl/nucleases completely removed the cellular component and supported the growth of fibroblasts and epithelial cells, while maintaining the integrity of the corneal epithelial BM [121]. Hashimoto et al describe a novel decellularization method using high-hydrostatic pressure (HHP) without detergents. The advantages of HHP include the elimination of bacteria and viruses, and the preservation of collagen that provides a relatively better mechanical support [125]. Similar to the HHP method, Huang et al reported a gentle physical decellularization method using supercritical carbon dioxide (SCCO2), with less processing time, lower cost and the elimination of pathogens to prepare an acellular porcine cornea. Compared to the Triton treatment, the use of SCCO2 enabled better removal of proteins with no immune response in a rabbit model at 6 months follow-up. Importantly, the maximum load of the DC is reduced, because of the increased spaces between the collagen fibrils due to the decreased GAG contents [126]. A final decellularization method worth noting employs pancreatic phospholipase A2 (PLA2) and SD. This approach preserves proteoglycans and the structure of the native corneal stroma, while removing most of the xenogenetic cells [119].
By preserving the corneal structure, the DC can provide excellent functionality with similar optical and mechanical properties to a native cornea with the capacity to support the confluent growth of the corneal epithelium, keratocytes, and endothelium, as well as the ability to degrade slowly while merging with the surrounding stroma during regeneration of the corneal epithelium and nerve growth [117, 122]. By co-culturing limbal epithelial cell-like (LEC-like) cells and corneal endothelial cell-like (CEC-like) cells derived from hESCs on the two sides of a decellularized porcine scaffold, a full-thickness artificial cornea substitute with similar thickness and mechanical properties to a native rabbit cornea was successfully constructed, with multilayered epithelium-like cells and a uniform monolayer of CEC-like cells observed. The thickness, endothelial cell density, and mechanical properties of the construct were similar to that of native rabbit corneas. Scaffolds could be integrated in a rabbit model with the restoration of transparency at an 8 week follow-up, indicating the DC to be a promising substrate for full-thickness corneal bioengineering [127].
3.5. Chitosan
Chitosan is an abundant naturally derived polymer that is synthesized by deacetylation of chitin. For corneal bioengineering, chitosan has primarily been employed as a substitute for AM as a cell carrier to reconstruct the ocular surface or endothelium. It has been reported that primary bovine corneal epithelial cells (BCECs) seeded on pure chitosan membrane showed improved proliferation and attached more quickly than those on AM. Moreover, pure chitosan membrane is able to preserve the phenotypes of BCECs similar to AM [128]. Furthermore, chitosan surface modification can have a positive effect on generating a monolayer of HCECs, while non-modified substrates cannot [129]. For endothelial regeneration, a chitosan membrane with controllable biodegradation rates can be achieved by incorporating polycaprolactone (PCL), a slow degrading synthetic polymer. BCECs cultured on this blended membrane showed normal appearance, and proliferated well, forming a continuous monolayer [130]. In addition to serving as a potential substrate for HCECs, the incorporation of chitosan can promote the optical transparency and mechanical strength of collagen membranes [78]. Because of the poor solubility of chitosan in physiological solvents, the research focus has been shifted to hydroxyethyl chitosan (HECTS), a water-soluble derivative of chitosan. With comparable optical transparency, water content, and higher glucose permeability compared to the natural human cornea [131, 132], membranes made up of hydroxypropyl chitosan, gelatin and chondroitin sulfate can improve the growth of HCECs and rCEN cells, promote the growth of primary rCEN cells and corneal epithelium restoration in vitro and are tolerated well in the anterior chamber of an in vivo rabbit model [131–133]. Although chitosan has demonstrated great potential for corneal bioengineering, it is more suitable to be utilized as blended, rather than pure scaffolds.
3.6. Thermal responsive polymers
Stimuli responsive polymers such as poly(N-isopropylacrylamide) (PNiPAAm) and PNiPAAm-based copolymers are being increasingly used in bioengineering, because of their thermoresponsive hydrophobic and hydrophilic properties, which enables the detachment of cell sheets without the use of enzymatic digestion. The ability to generate carrier-free cell sheets makes it a promising technique for corneal epithelium and endothelium engineering. While several publications support the use of both corneal epithelium [134] and endothelium cell sheets [135] derived from temperature-responsive substrates, endothelium construction has attracted the most interest. Using rabbit limbal epithelial stem cells isolated from rabbit ocular surface, corneal epithelial sheets with compact multilayered cell sheet architecture were successfully generated from PNiPAAm grafted cell culture surfaces using poly(vinylidene difluoride) (PVDF) membrane and applied for autologous transplantation. The cell sheets attached to an exposed rabbit corneal stroma within minutes without suturing and successfully reconstructed the corneal epithelium [134]. The structure and function of HCEN cell sheets detached from pure PNiPAAm are able to restore corneal clarity without causing inflammation or rejection in rabbit models [135, 136]. The activity of Na+, K+-ATPase and existence of pump sites were confirmed in vitro and more importantly, proper stromal hydration was found and corneal clarity was gradually restored in the rabbit model [135, 136]. Apart from pure PNiPAAm, copolymerization with glycidylmethacrylate (NGMA) allows the manipulation of the critical temperature and comprises epoxy groups for the incorporation of biomolecules. The use of a copolymer of N-isopropylacrlamide (NiPAAm) and diethyleneglycol methacrylate (DEGMA) enables faster cell detachment with a critical temperature, closer to the physiological range [137, 138]. Teichmann et al reported a thermal responsive carrier using poly(vinyl methyl ether) (PVME) where layer thickness, stiffness, switching amplitude and bioactive modification can be coordinated to regulate the initial immortalized HCEN cell adhesion and detachment of cell sheets [139]. Despite all the advantages, given the thermally detached cell monolayers are usually fragile, the use of compatible delivery methods is important for the surgical manipulation of sheet grafts. Varied carriers have thus been investigated for the delivery of constructed epithelial (PVDF) [134] and endothelial layers (stroma bed and gelatin) [135, 136].
3.7. Other synthetic polymer hydrogels
A variety of synthetic materials including poly(vinyl alcohol) (PVA) [140], poly(2-hydroxyethyl methacrylate) (PHEMA) [141], polyethylene (glycol) Diacrylate (PEGDA) [142, 143], poly(lactic-co-glycolic acid) (PLGA) [144] and poly(ethylene glycol)/poly(acrylic acid) (PEG/PAA)-based hydrogels [145–147] have also been applied in corneal bioengineering, as either substrates for corneal cells or artificial cornea substitutes. While exhibiting excellent permeability, controllable mechanical and optical properties, all of these materials display poor cell-adhesive function without surface modification [148]. For example, to improve the attachment and growth of primary rabbit and human corneal cells, electrospun PVA, PHEMA hydrogel and PEGDA hydrogel were modified with collagen type I, phosphate groups and RGD, respectively [140, 141, 149]. Moreover, hydrogels functionalized with amine can support epithelization using HCECs [148]. Similarly, collagen type I coated PLGA films can better serve as substratum for rCEN cells [144]. An artificial cornea fabricated using ZnS/PHEMA/PAA hydrogel demonstrated high refractive index [145] and PEG/PAA hydrogel showed high glucose permeability, while remaining optically clear in situ for up to 2 weeks [146, 147]. Recently, poly(glycerol sebacate) (PGS), a novel biodegradable elastomer, was cured into films and capable of maintaining the viability of HCECs. However, there have been no reports of the mechanical or optical properties, or in vivo testing to date [150].
4. Material surface modification
4.1. Topographical modification
The influence of topographical cues and surface chemical modification are important for cell support regardless of the tissue type being engineered [151, 152]. For topographical modification, the most commonly used technique to create varied patterned features is soft lithography [101, 116, 151, 153–158]. Briefly, feature-patterned master molds (for instance, patterned silicon substrates), are covered with PDMS, cured on the patterned substrate and then removed to be used as patterned templates for cell seeding. Anatomically, for corneal epithelial engineering, the key findings are that HCEC contact guidance is not only modulated by the scale of the topographical cues [151] and HCLE cells seeded on patterned silk created by lithographic techniques but suggest parallel line patterned surfaces promote initial cell attachment by 36%–54% and more that twofold increase in focal adhesion localization [153]. In addition, the adhesion and proliferation rate of HCECs on collagen film can be promoted by inducing a micro-rough surface using a freeze-drying technique [154]. Interestingly, for corneal stromal bioengineering, whilst both the depth and width of the grooves influence HCFs alignment, the former appears to play a more critical role for cell orientation [101]. On poly (L,D lactic acid) (PLDLA) electrospun nanofibrous substrates, HCS cell orientation and differentiation can be directed, with stiff and orthogonally arranged nanofiber layers resulting in keratocyte-like morphology other than those of HCFs [155]. Nara et al employed direct-write assembly and prepared parallel patterned thermos-responsive substrates that can guide goat corneal keratocyte alignment. However, these patterned substrates could not produce complete sheet restoration [157]. To compare the effect of different topography on endothelial cells, various kinds of patterns including channels, gratings, concentric circles, wells and pillars have been studied. Bovine corneal endothelial cells grown on pillars provide a lower coefficient of variation of area, enhanced Na+/K+-ATPase activity and a density of microvilli similar to the native cornea compared to other patterns [156]. A similar finding on HCECs reported by Rizwan et al showed that higher Na+/K+-ATPase expression was found on the substrates of square-array and hexagonal-array of 1 mm pillars with 6 mm spacing compared to flat substrate. Specifically, square-array topography increased ZO-1 and Na+/K+-ATPase expression higher, while hexagonal-array induced more favorable cell morphometry such as high cell densities, low cell areas and homogenous cell size, thus improving the quality of the regenerated HCEC monolayer [116].
Apart from stereo lithography, the effect of topographical modification on corneal cell growth and subsequently corneal tissue regeneration can be explored via a noteworthy fabrication technique, namely electrospinning. Electrospinning is a widely investigated fiber fabrication method that applies a voltage between a syringe and a collector to draw fibers from polymer solutions dispensed by the syringe [159]. Electrospun scaffolds are increasingly being investigated to develop biomimetic bioengineered corneas as a result of the bionic reconstruction of ECM, which showed the promotion of corneal cell adhesion, proliferation, migration and differentiation [129, 160]. In many instances natural materials are used including SF [94], gelatin [161], collagen [129], HA [129]) and synthetic polymers PCL [162], PLGA [163], RGD (PEUU) [164], poly-L/D-lactide (PLDLA) [155], poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) [162], with the latter characterized by outstanding mechanical properties. While the use of natural materials is limited by inferior mechanical toughness and therefore handling for surgery, synthetic polymers such as FDA approved PLA, PLGA, and PCL have been employed extensively. The application of electrospun scaffolds is primarily limited to serve as substrates for the growth and delivery of corneal cells and the exploration of cell-substrate interactions. Electrospun substrate as a replacement for AM to support the regeneration and delivery of corneal epithelium has been widely investigated. For instance, (PHBV)/gelatin [160], collagen/chitosan/hyaluronate(HA)/PEO [129], PLGA [163] electrospun substrates have been shown to be suitable substitutes for AM. Gelatin/PLLA-based substrates have been used to investigate the relationship between HCECs and keratocyte cell behaviors and fiber arrangement [165]. HA/PEO electrospun film was characterized with excellent mechanical properties, and comparable optical and biological performance in vivo for rabbit ocular restoration [129]. Ortega et al electrospun biodegradable PLGA fibers onto PEGDA ring with microfeatures to mimic corneal limbus with the rabbit corneal limbal epithelial cells well attached and proliferated on the designed microstructure [159]. Furthermore, PCL electrospun nanofibers support HCLE cell phenotype to facilitate cornea epithelium construction and promote the cell attachment of rabbit LSCs and rabbit keratocytes [162]. Notably, PLGA electrospun scaffold is not only biocompatible with HCS cells but capable of maintaining a keratocyte phenotype [155] and multilayered rabbit limbal epithelial cells [163]. Apart from the choice of material, the ease of manipulating mechanical properties, transparency and biological properties is also determined by polymer components, special arrangement and functionalization [165–167]. For example, the degree of alignment of fibers effects the mechanical properties of scaffolds [165]. Aligned fibers mirror the natural configuration necessary for cell support and differentiation [161, 168]. For instance, HCS cells cultured on electrospun PEUU aligned scaffolds tend to secrete collagenous matrix with similar alignment to natural corneal stroma [164]. Similar results have shown that aligned electrospun collagen scaffolds tend to dedifferentiate rabbit corneal fibroblasts from the myofibroblast phenotype [167]. Salehi et al reported electrospun nanofibrous PGS/PCL semitransparent substrate that is able to support the growth of HCEN cells with hexagonal morphology and direct human conjunctival epithelial cells growing along the aligned fibers with the incorporation of PGS improve the cell proliferation and viability [169]. Earlier studies demonstrated the potential of electrospun scaffolds in corneal tissue formation as a result of the favorable mechanical properties, transparency and the biocompatibility with various corneal cells. However, few in vivo studies have been performed to date.
4.2. Chemical modification
Modifications with proteins, peptide and chemical groups are commonly utilized to promote corneal cell growth. Apart from the widespread modification of RGD or collagen on synthetic polymers, carbon chain length of amine modifications are able to impact the attachment of epithelial cells and stromal cells [148]. A study of collagen type I, collagen type IV, fibronectin (FN), FNC coating mix and laminin modifications indicate all these coatings facilitate cell adhesion, with FN coating being the most favorable for HCEN cell growth and optimum compact cellular morphology [170]. Interestingly, chitosan coating was also found to improve the growth of HCEC line [129].
Peptide amphiphiles (PAs) are peptide-based molecules typically composed of hydrophilic and hydrophobic domains and of great interest in tissue engineering by virtue of their ability to self-assemble into high-aspect-ratio and highly bioactive nanostructures under certain physical conditions (pH, temperature and ionic strength) [171]. PA generally comprises three components: a hydrophobic tail, a beta-sheet forming amino acid, and a hydrophilic head that promotes water-solubility and biocompatibility. Self-assembly of PA can be guided by the hydrophobic collapse of the tail and achieved by hydrogen bonding between beta-sheet forming amino acids (figure 6). More specifically, hydrophobic tails of PAs were internalized and the hydrophilic heads exposed as bioactive interacting surfaces [172]. For tissue engineering, PAs are commonly applied in different formats, including coating, solution and hydrogel formats. For corneal bioengineering, PA has primarily been used for coating or in solution due to the inferior mechanical toughness of hydrogels, able to support a wide range of corneal cells [171]. As previously mentioned, RGD has been widely coated on various biomaterials to promote corneal cell interaction and adhesion. In this context, Gouveia at al employed PA consisting of RGD-containing peptides as a film coating that was able to enhance the adhesion, proliferation and alignment of HCFs and enable the construction of 3D lamellar-like stromal tissue [173]. By also adding a matrix metalloprotease (MMP)-cleavable sequence, the PA not only acted as a substrate for the attachment and growth of HCS cells in a quiescent phenotype but enabled self-release after adding retinotic acid to the culture medium to facilitate the detachment of regenerated stromal matrix from the base [174]. In addition, Jones et al [175] report a special PA: C16-KTTKS, which self-assembled into nanotape structures being able to stimulate collagen I production from HCS cells. Finally, Uzunalli et al [176] applied a laminin-derived sequence YIGSR into PA to regulate cell adhesion for corneal stroma regeneration. Compared to RGD-PA, as-prepared PA demonstrated enhanced cell proliferation, keratocyte migration, and collagen I synthesis both in vitro and in vivo.

Figure 6. Examples of supramolecular self-assembled PA nanostructures: (A) nanofibers; (B) micelles; and (C) multilayered nanotapes. All three structures have a hydrophilic outer corona consisting of bioactive peptide (blue), self-assembly-inducing/spacer sequence (white), and a hydrophobic inner core with organized and/or non-organized PA tails (red and green, respectively). Reprinted from [171] CC BY 4.0.
Download figure:
Standard image High-resolution image5. In vivo performance
Due to the excellent intrinsic biocompatibility, collagen gel, both pure and blended developed by various kinds of methods, have shown promising performances in vivo. From 24 months follow-up of a phase 1 clinical study on the EDC crosslinked collagen gel scaffold, the fabricated implants remain stable and avascular, with corneal re-epithelialisation, restored tear film, recruited stromal cells, and nerve regeneration [177]. Varied hybrid collagen gels, such as collagen/chitosan [63], glutaraldehyde crosslinked collagen/TERP [75] were well tolerated in pig corneas for 12 months and dogs for 16 weeks, respectively. Being applied as full-thickness implants in guinea pigs, collagen–MPC hydrogels integrated into host corneas and promoted regeneration of corneal tissue and nerves. Electrical activity recorded from the regenerating corneal nerves suggested that the regeneration of corneal nerves into the implant started at 3 months post-surgery and the nerves were functionally active within the implant by 8 months [77].
Although surface modifications are normally necessary to promote corneal cell growth, silk-based films have demonstrated favorable in vivo responses. RGD-functionalized five layers of stacked silk implanted in the lamellar pocket at the limbus in the rabbit models can maintain their integrity and transparency over 180 days without causing immunogenic and neovascular responses or degradation of the rabbit corneal stroma. The regenerated ECM deposited and adhered well to the surface of the silk films [105]. Porous silk films prepared using PEG were inserted into stromal pockets at the limbus of rabbits and were gradually replaced by corneal stromal tissue, while stromal cells infiltrated into the silk films without causing inflammatory reaction 6 months later. Notably, unlike porous silk films, non-porous ones did not degrade at 6 months [93]. Transparent blended films with aloe vera gel and SF as a carrier for corneal endothelial cells was developed by Kim et al by seeding rCECs on the films to reach confluency. Transplanted into the anterior chamber of the rabbits' eyes together, the constructs attached well to the surface of the corneal stroma and 4 weeks later, started to integrate with the surrounding corneal tissue with the rCECs firmly adhered and functional. Moreover, rabbit cornea transplanted using hybrid AV/SF scaffolds with rCECs revealed a higher corneal transparency compared to that of pure SF films with rCECs, indicating the improvement of clarity brought by the AV. However, many constructs were difficult to insert during the transplantation surgery, because of insufficient mechanical properties [178]. An investigation on how transplantation techniques affect the biological responses of silk films in rabbits is stimulating and worthwhile noting. Porous silk films transplanted into different locations of the ocular surface (peripheral-median P-M, central-superficial C-S, central-deep C-D) were used to assess the in vivo response. The in vivo response of the films was found to be dependent on the method to create the corneal pocket, the position of the sample and sutures. To be more specific, with the use of flushing air (instead of using a surgical a blade), the C-D position (compared to P-M and C-S) and a lack of sutures resulted in less injury and inflammatory responses. Two months after the surgery, a comparison of the degradation of silk films with low, medium and high beta-sheet (crystalline) content showed that low beta-sheet samples lost structural integrity, while medium and high beta-sheet content films remained at the initial stage [179]. Although fibroin film is designed as a transplantable carrier, improvement in physical properties is needed to graft the sheet onto a curved surface such as the cornea.
One common application of gelatin for corneal bioengineering is to serve as carriers for endothelial regeneration and delivery. For instance, EDC crosslinked intact and porous gelatin carriers have been inserted into the anterior chamber of rabbit eyes and no change was observed in the cell morphology of the rCECs by day 3 [112, 115]. Being implanted into the anterior chamber of the endothelium-removed rabbit model, EDC crosslinked heparin-modified gelatin remained transparent by week 3 with little surrounding intraocular inflammation [114]. Glutaraldehyde crosslinked gelatin seeded with rabbit fibroblast and fibroblast precursors were transplanted into rabbit eyes with lamellar dissection. While both carriers were well tolerated during the 4 weeks follow-up, the gelatin/precursor group showed more intense expression of ECM molecules [111]. Longer-term observations of the biological responses of gelatin used in vivo are however limited.
Due to the well-maintained integrity of ECM, DC has demonstrated excellent biocompatibility in vivo. Porcine corneas, decellularized using Tris-HCL [117], SDS [118], PLA2/SD [119], HHP method [125], SCCO2 [126], have been transplanted into rabbit corneal stroma pockets to investigate the biological responses. The implants were normally capable of becoming completely transparent within 2 weeks, being maintained for at least 12 months after transplantation with the implants being able to support a stratified epithelium with infiltrated keratocytes. Zhang et al developed a full-thickness cornea substitute using LEC-like and CEC-like cells derived from human embryonic stem cells and decellularized bovine corneas. Being transplanted into a rabbit model using PK, no significantly high intraocular pressure was found in the follow-up period. By 8 weeks, the substitutes were well integrated with corneal tissue. LEC-like cells were completely replaced by host cells and the endothelial cell density of tumor endothelial cells was similar to that of native rabbit corneas. Compared with a decellularized bovine cornea without cells, the full-thickness graft showed less neovascularization with new blood vessels only in the periphery, while its counterpart was completed invaded [122].
Research relating to the in vivo performance of chitosan is limited. It is reported that after HECTS/CS blended membranes were transplanted into the anterior chamber of rabbits, the cornea still retained clarity and no apparent symptoms could be detected [132]. For corneal epithelial bioengineering, rCEC-seeded chitosan membranes were transplanted onto the ocular surface in rabbits, promoting corneal recovery [133] and that chitosan coatings were able to promote corneal epithelial recovery in a rabbit corneal defect model [129].
While most synthetic hydrogels are applied as substrates to investigate fundamental cell behaviors, most of them did not receive in vivo evaluation. As an artificial cornea, PEG/PAA hydrogel was able to remain optically clear and show excellent tolerance after being sutured into the stroma of a rabbit for 2 weeks [146]. A comparison between poly(ethylene glycol) diacrylate (PEGDA) and poly(ethylene glycol) diacrylamide (PEG-dAAm) hydrogel showed that in terms of in vivo biocompatibility in rabbits, the PEG-dAAm-based hydrogel was superior to the PEGDA-based hydrogel, with far longer maintenance of corneal clarity up to 6 months, while the PEGDA-based gel implanted ones showed haze and opacity within 10 weeks [147].
6. The importance of cell biology
Considering the different cell types throughout the cornea and the likely requirement of more than one type of cell support substrate with a composite of biomaterials, there is a need for extensive optimization studies towards merging the components for a complete and functional cornea. For example, electrospun aligned PEUU substrate [164], combined with growth factors [180] and the selection between hCSSCs and HCFs will result in different collagen fibril matrices and therefore different tissue constructs. Specifically, multilayered lamellae with orthogonally oriented collagen fibrils that accurately mimic the HCS tissue will likely require the combination of fibroblast growth factor (FGF-2) and transforming growth factor-beta3 (TGF-b3). Both of these growth factors can stimulate the secretion of collagen from cells resulting in the directional growth of collagen fibrils and orthogonal fibril orientation of a collagenous layer [180]. Regarding cell choice, hCSSCs should differentiate into keratocytes and secrete multilayered lamellae with well-organized collagen fibrils, while HCF cells are more likely to differentiate into fibroblasts and generate a less-organized collagen construct [181]. Similar results can be expected for porous gelatin hydrogel-based constructs, with fibroblast precursors likely better candidates for corneal stroma engineering compared with fibroblasts, because of higher collagen I expression in vivo after implantation [111]. Moreover, the use of serum tends to induce hCSSCs to become fibroblastic. In contrast, a keratocyte-like morphology arises in the absence of serum [155]. Compared to culturing HCECs and stromal cells alone, co-culturing both cells demonstrated improved differentiation and growth for both of the cells [182]. A study on the influence of serum concentration, oxygen tension for incubation and macromolecules (carrageenan)-supplemented media on ECM deposition in HCF culture interestingly found that the low oxygen tension coupled with supplemented media in the culture significantly promoted ECM deposition at day 14 [183].
Practically, the use of seeding cells to differentiate or modulate the local tissue environment thereby promoting regeneration and repair remains a logical progression for research. A breakthrough on corneal epithelial regeneration using tissue-engineered cell sheets composed of autologous oral mucosal epithelium offered a new treatment for ocular trauma or disease. After scar tissues on the ocular surface were removed, cell sheets constructed by culturing oral mucosal tissue from patients on temperature-responsive polymer poly(N-isopropylacrylamide) were transplanted into the patients' eyes. Excitingly, complete re-epithelialization was detected within 1 week in all treated eyes and corneal transparency was restored and well maintained through long-term follow-up (mean period of 14 months) [184]. Yang et al previously showed that human amniotic epithelial cells could differentiate into conjunctival epithelium-like cells and goblet cells with partially physiological function thereby providing successful restoration of ocular surface integrity in a rabbit model of conjunctival disease [185]. More directly, Mathan et al successfully isolated and transplanted sphere-forming cells from the peripheral cornea for the potential use of targeting in vivo ocular surface regeneration and stem cell repopulation [186]. Yet challenges remain. Understanding the detailed mechanism of differentiation induced by the niche, the method of immortalization of differentiated cells and the physiological functions of tissue-engineered surfaces is essential to long-term success. The microenvironment appears to play an important role in the directional differentiation and fate of the implanted cells [187]. As evidence, Mathan and co-authors found that corneal repopulation differed from scleral cell repopulation. The authors hypothesized that inherent characteristics within cells may lead to preferential changes (i.e. a cell derived from the limbus would like to reform a limbus or limbal niche). Separately, the authors found sphere implantation indicated a similar reactive biological response to the wounding process. Regardless, current findings provide tangible evidence to support long-held theoretical beliefs. The concurrent use of biomaterials may provide additional benefits and further research is essential to optimizing an effective approach to corneal tissue regeneration.
7. Conclusion and future challenges
Over the past few years, many different types of bioengineered substitutes have been developed to address the shortage of qualified corneal donors. Biomaterials used are largely limited to natural polymers able to support cell growth and function in vitro, and tissue regeneration in vivo. Collagen remains the best candidate in part due to its excellent biocompatibility from intrinsic RGD, with the main challenges in the biomimetic fabrication of aligned collagen fibrils mirroring corneal stromal inner structure. Silk has shown excellent optical clarity and more importantly, controllable degradation rates and mechanical properties, unlike other natural polymers. In terms of pursuing cell-biomaterial interactions, the ease of patterning and chemical modification enables silk to outweigh other materials. Derived from collagen, gelatin does not show any better overall performance. Alternatively, chitosan is easily functionalized and highly biodegradable and could be more useful when acting as an incorporated component in blended scaffolds. Despite their excellent physiological properties, the modification of synthetic polymers (e.g. RGD, collagen addition) is required to achieve satisfactory biological performance. Notwithstanding, they have significant potential as artificial corneas or substrates for various kinds of corneal cells, with most of the candidates mentioned above showing favorable ability to support the growth and delivery of diverse cell types.
In spite of the progress and promise of a bioengineered cornea, there are many challenges to overcome. Firstly, human trials are necessary to definitively determine the efficacy of corneal substitutes. Currently, clinical trials of bioengineered corneal substitutes are by-and-large limited to collagen-based materials with other materials rarely reported [177, 188]. A 12 month follow-up of LSC therapy and long-term corneal regeneration conducted on 113 eyes of 112 patients with burn related LSC deficiency showed that the total success rate by one year, as characterized by a transparent, avascular, and stable corneal surface being restored, was 76.6% [189]. A significant finding is that a minimum number of 3000 p63-bright holoclone-forming LSCs is essential and suggested for clinical success of autologous LSC transplantation [189]. Most bioengineered substitutes are merely designed to fulfill the requirements of transparency, biocompatibility and biodegradability, with other important properties such as innervation and regeneration often overlooked. Corneal innervation is critical to corneal sensation and protection, with optimal nerve density important for wound healing. Notwithstanding, some studies have demonstrated the reinnervation of collagen-based grafts in vivo [66, 74–77, 177], but reports on neural regrowth of other materials remain limited. Notably, Wang et al recently developed an innervated silk-based multilayered corneal epithelium and stroma including innervation induced and directed by nerve growth factor in vitro. Increased IVL, GJA4, ALDH3A1 and KERA expression of innervated models indicated better corneal epithelial maturity and corneal stromal transparency compared to non-innervated ones, which further demonstrated the key role of innervation in the maintenance of the corneal epithelium and stroma (figure 7) [190]. Then, while a great number of studies have reported the effect of the characteristics of scaffolds (composition, structure, pattern, etc) on in vitro response, few were carried out on the influence of those on the in vivo response, which might be more significant when pursuing the best substitute for corneal tissue. The use of a porous structure [101] or combining propionamide [106], for instance, tends to induce a faster biodegradation rate in an animal model. Finally, recognition of the structural distinction of different cornea tissue layers has underscored the need for a variety of strategies to develop substitutes amenable to generating each layer. Despite the challenges, progress is being made towards identifying efficacious and accessible substrates capable of supporting cell growth and delivery, and tissue restoration towards recapitulating bona fide corneal tissue. Most challenging, has always been the fabrication of a stroma consisting of parallel lamellar sheets and a highly ordered microstructure of collagen fibrils with functional keratocytes and proteins to mimic the cornea stroma, as a result of the anatomical and functional importance of the stroma. Once it is achieved, bioengineering a whole-thickness cornea should be much easier, since substantial progress has been claimed on the regeneration of the cornea epithelium and endothelium, which are mainly cell layers. Engineering corneal substitutes with all layers for full-thickness PKP procedure would be preferable. Currently, protocols to fabricate a whole-thickness cornea are cell-free and cell-encapsulated membranes. Recently, electrochemically compacted collagen matrices with improved strength and modulus after being crosslinked by EDC/NHS or genipin demonstrated highly dense structure and comparable transparency to the native cornea, suggesting great potential for corneal tissue engineering [191]. Apart from collagen gel, human keratocytes encapsulated into fibrin-agarose scaffolds with similar transparency, and UV absorption characteristics to the native human cornea [192, 193], have been built into a whole cornea by seeding corneal epithelial cells and endothelial cells on each side [194]. Zhang et al recently developed a biomimetic 3D corneal model by loading patterned silk films and collagen gel onto a dome-shaped post and more interestingly, the effect of mechanical strain on human keratocyte phenotype and ECM formation was investigated (figure 8). The keratocyte alignment and ECM arrangement were directed by the topography of the silk films and compared with traditional 2D culture, and higher expression of keratocyte marker and expression of ECM were detected in the 3D model. Moreover, higher expression of keratocyte marker was also observed on a 3% dome-shaped mechanical strain cornea model compared with flat-shaped strain [158]. Gouveia et al successfully fabricated auto-generated scaffold-free 3D corneal equivalents by stacking HCS cell sheets collected from PA coatings and subsequently culturing the multilayered cell sheets for 21 days. The corneal equivalent was not only transparent but well integrated into rabbit models after 9 months. Other interesting findings included the alignment of stromal cells and secreted collagen fibrils were directed by the topography of growth substrates with different anisotropies, with tissues comprising aligned collagen fibrils tending to be thicker, denser and more resistant to proteolytic degradation compared to those comprising randomly oriented fibrils. This work is definitely significant for fabricating a scaffold-free human tissue that mimics the corneal stroma with controlled structure and functional properties [195].

Figure 7. Schematics of human cornea and an in vitro 3D corneal tissue model. Scale bars = 3 mm. Reprinted from [190], copyright (2017), with permission from Elsevier.
Download figure:
Standard image High-resolution image
{kind=link}
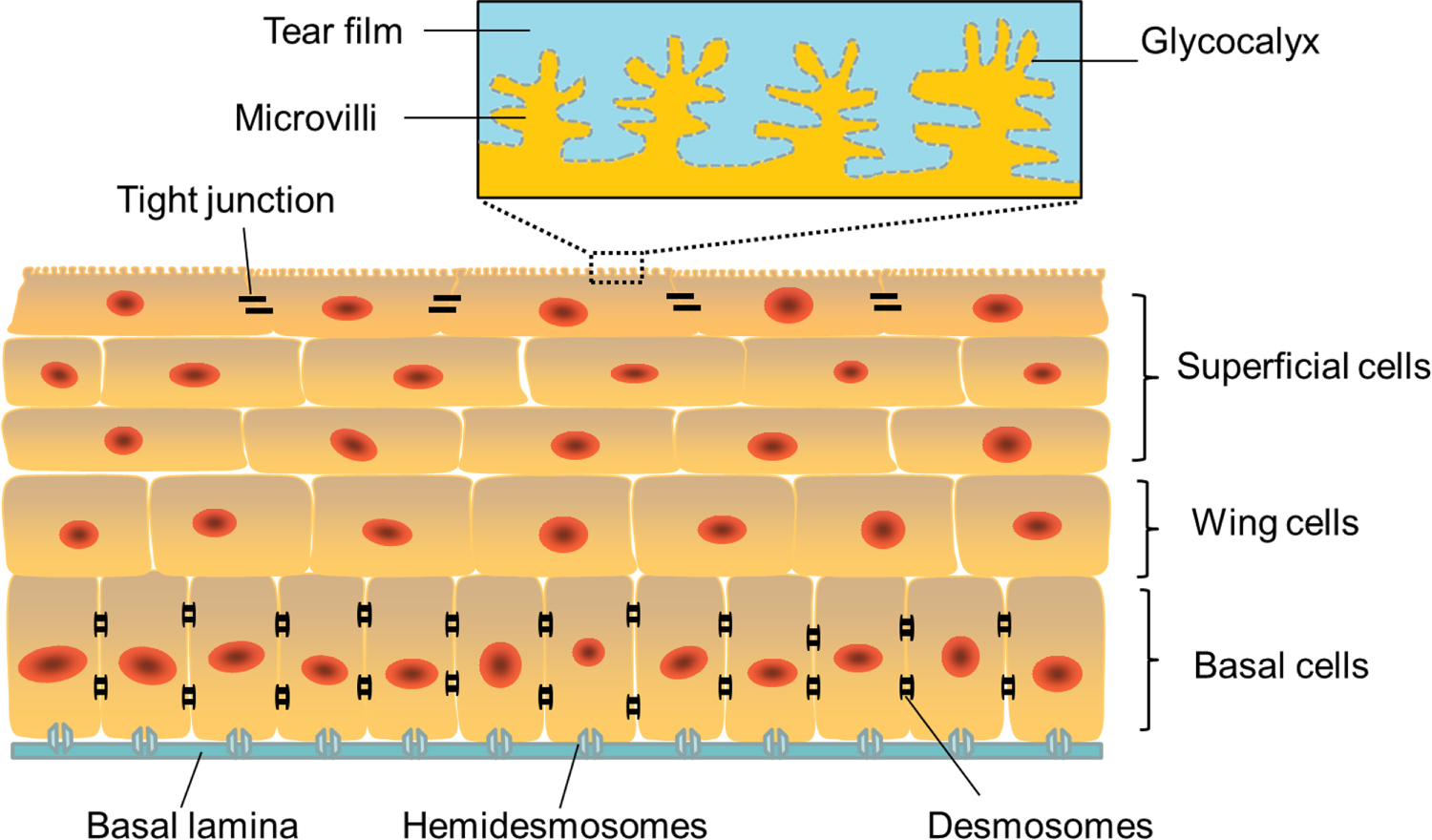{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 8. The synergistic effects of topography and mechanical strain on keratocyte behavior in a biomimetic 3D corneal model. (A) Schematic for the biomimetic 3D corneal model using patterned (600 grooves mm–1) SF films, collagen I hydrogel, and 3% dome-shaped strain. Reprinted from [158], copyright (2017), with permission from John Wiley and Sons.
Download figure:
Standard image High-resolution image{kind=link}
In spite of the challenges, there is little doubt that corneal implants will be partly or completely engineered in vitro as qualified alternatives to current treatments, thereby addressing the present shortfall in donor corneas. The rapid advancement in biomaterials, cell biology and tissue engineering methods such as bioprinting will facilitate the formation of durable biomimetic structures that are customized to the individual needs of recipients, improving transplant durability and functionality, and ultimately the quality of life for the large numbers of patients worldwide suffering from corneal blindness.
Acknowledgments
The authors wish to acknowledge funding from the Australian Research Council (ARC) Centre of Excellence Scheme (CE140100012). Professor Gordon Wallace acknowledges the support of the ARC through an ARC Laureate Fellowship (FL110100196).