
Abstract
Quadrupeds achieve rapid and highly adaptive locomotion owing to the coordination between their legs and other body parts such as their trunk, head, and tail, i.e. body–limb coordination. Therefore, a better understanding of the mechanism underlying body–limb coordination could provide informative insights into the improvement of legged robot mobility. Sprawling locomotion is a walking gait with lateral bending exhibited in primitive legged vertebrates such as salamanders and newts. Because primitive animals are anticipated to possess the essence of quadruped motor control, their locomotion helps better understand body–limb coordination mechanisms. Previous studies modeled neural networks in salamanders and employed it to control robots and investigate and emulate sprawling locomotion. However, these models predefined the relationship between the legs and the trunk, such that how body–limb coordination is attained is largely unknown. In this article, we demonstrate that sensory feedback facilitates body–limb coordination in sprawling locomotion and improves mobility through mathematical modeling and robot simulations. Our proposed model has cross-coupled sensory feedback, that is, bidirectional feedback from body to limb and limb to body, which leads to an appropriate relationship between the legs and the trunk without any predefined relationship. Resulting gaits are similar to the sprawling locomotion of salamanders and achieve high speed and energy efficiency that are at the same level as those of a neural network model, such as conventional models, optimizing the relationship between the legs and the trunk. Furthermore, sensory feedback contributes to the adaptability toward leg failure, and the bidirectionality of feedback facilitates parameter tuning for stable locomotion. These results suggest that cross-coupled sensory feedback facilitates sprawling locomotion and potentially plays an important role in the body–limb coordination mechanism.
Export citation and abstract BibTeX RIS
1. Introduction
Quadrupeds move in their natural habitat without any difficulties. Such agile locomotion is achieved by the coordination between their legs and other body parts, such as their trunk, head, and tail, i.e. body–limb coordination [1–5]. For example, galloping cheetahs bend their bodies to elongate stride length and to kick the ground firmly, and walking lizards swing their tails to achieve balance [1, 2]. These behaviors suggest that body–limb coordination has an essential role in quadruped motor control. However, the mechanism responsible for body–limb coordination is largely unclear. A better understanding of this mechanism would contribute to the design of agile quadruped robots in addition to elucidating the motor control in quadruped animals.
Sprawling locomotion is a remarkable behavior that helps understanding the body–limb coordination mechanism; it is a walking gait with lateral bending and is adopted by many reptiles and amphibians [2–5]. Although they have short legs, lateral bending facilitates longer stride and stronger thrust during locomotion [3]. Moreover, the behavior is exhibited in primitive legged vertebrate such as salamanders and newts [5, 6], such that an essence of body–limb coordination might underlie the locomotion mode.
Sprawling locomotion is controlled by a distributed neural network, called central pattern generator (CPG) with sensory feedback from peripheral nerves according to decerebrate salamander experiments [7]. Based on the findings, several salamander robots have been developed to emulate and investigate sprawling locomotion [5, 6, 8–10]. These studies modeled distributed networks in coupled oscillators and employed them for robot control. Most of the models predefined the relationship between the leg and the trunk motion by the neural-coupling of oscillators. Ijspeert et al presented a simple spinal cord model and implemented it in an amphibian robot that replicated sprawling locomotion and anguilliform swimming in response to the strength of a descending command [5, 6]. Zhong et al developed a salamander robot that succeeded in moving over complex terrains, and they observed that lateral bending improves forward, rotational, and lateral locomotion [8]. Yin et al proposed a CPG model with non-linear oscillators that facilitated coordination between the leg and the body to increase step length [9]. In contrast to these centrally defined body–limb coordination models, Horvat et al presented a spine–limb coordination controller with sensory feedback and demonstrated sprawling locomotion [10]. Although the study presented the sprawling locomotion model with sensory feedback, the extent to which sensory feedback is involved in shaping body–limb coordination was not extensively investigated. Therefore, studying the role of sensory feedback in sprawling locomotion is an important step to understand fundamental principles of body–limb coordination.
In this article, we demonstrated several utilities of sensory feedback in sprawling locomotion through mathematical modeling and robot simulations. Our proposed model has cross-coupled sensory feedback, bidirectional feedback from body to limb and limb to body, which helps establish appropriate body–limb coordination immediately without relying on a predefined relationship of neural-couplings. The feedback utilizes peripheral proprioceptive or exteroceptive sensory information such as position sense and pressure sense. This sort of sensory information including physical and environmental conditions helps improve the traveling ability. Simulation studies investigated the locomotion speed, cost of transport (CoT), and fault tolerance on flat terrain. Resulting gaits are similar to the sprawling locomotion of salamanders and achieved mobility that is at the same level as that of a neural network model, such as conventional models, optimizing the relationship between the legs and the trunk. Furthermore, the sensory feedback model exhibited higher adaptability toward hind legs failure than the neural network model. The bidirectionality of feedback also contributes to superior mobility compared with unidirectional feedback from body to limb or from limb to body or compared with those models without feedback, and stable locomotion can be achieved without precise parameter tuning. These results suggest that cross-coupled sensory feedback facilitates sprawling locomotion and potentially plays an important role in body–limb coordination mechanisms.
The remainder of this article is structured as follows: section 2 describes a decentralized control for body–limb coordination and the details of cross-coupled sensory feedback. Section 3 describes the details of the simulation experiment and the results. Section 4 discusses the usefulness of sensory feedback in sprawling locomotion and the contribution of feedback bidirectionality. In section 5, the conclusions and recommendations for future studies are presented.
2. Model
Upon investigating the extent to which sensory feedback is involved in body–limb coordination mechanism, we considered four feedback rules as follows (figure 1):
- (i)Sensory feedback from limb to limb
- (ii)Sensory feedback from body to limb
- (iii)Sensory feedback from limb to body
- (iv)Sensory feedback from body to body.
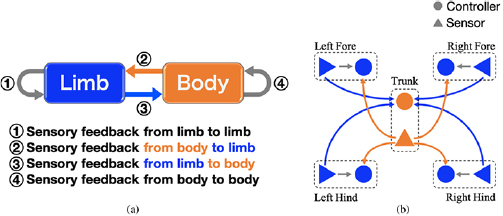
Figure 1. Overview of cross-coupled sensory feedback, (a) four sensory feedback rules that facilitate adaptive legged locomotion and (b) feedback structure of proposed robot model. The model uses force sensors at the feet and a torque sensor at the trunk for the cross-coupled feedback.
Download figure:
Standard image High-resolution imageWe presume that the interplay of four rules facilitates adaptive locomotion, particularly the cross-coupled feedback comprising second and third rules, bidirectional feedback from body to limb and from limb to body, and establishes self-organized body–limb coordination without any predefined relationship between the legs and the trunk. The first rule coordinates four legs to locomote, while counteracting gravity. The second and third rules are the cross-coupled sensory feedback that generates adaptive body–limb coordination. The fourth rule orchestrates other body parts such as the trunk, head, and tail.
Here, we propose a simple robot model whose feedback structure is shown in figure 1(b). To focus on understanding body–limb coordination, we adopted the minimal mechanical system for sprawling locomotion, and because of the simplicity of the mechanical structure, omitted the fourth rule that orchestrates axial degrees of freedom. The following section describes the mechanical and control systems.
2.1. Mechanical system
Figure 2 shows an overview of the mechanical system comprising the trunk and four leg segments. The trunk segment has a rotary actuator and passive spring damper in the yaw direction. Each leg segment has two rotary actuators in the yaw and roll directions along with a phase oscillator that defines the target angles of the actuators:
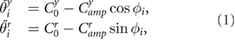
where  and
and  denote the target angles,
denote the target angles,  and
and  represent the neutral angles,
represent the neutral angles,  and
and  represent the amplitude of the yaw and roll actuators, respectively (figures 2(b) and (c)).
represent the amplitude of the yaw and roll actuators, respectively (figures 2(b) and (c)).  is the phase of the oscillator. When
is the phase of the oscillator. When  , the limb is in the swing phase; otherwise it is in the stance phase. The suffix i denotes the leg identifier (1:left fore, 2:right fore, 3:left hind, 4:right hind, respectively).
, the limb is in the swing phase; otherwise it is in the stance phase. The suffix i denotes the leg identifier (1:left fore, 2:right fore, 3:left hind, 4:right hind, respectively).
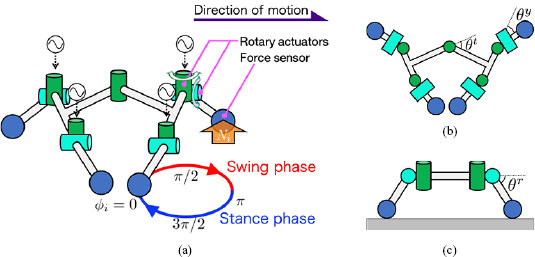
Figure 2. Overview of the mechanical system. The trunk has a rotary actuator and each leg has two rotary actuators that are controlled by the phase of the oscillators  . Force sensors positioned at the feet detect the ground reaction force Ni. Variables
. Force sensors positioned at the feet detect the ground reaction force Ni. Variables  ,
,  , and
, and  are angles of trunk-joint and leg actuators in the yaw and roll directions, respectively. (a) Oblique view. (b) Top view. (c) Front view.
are angles of trunk-joint and leg actuators in the yaw and roll directions, respectively. (a) Oblique view. (b) Top view. (c) Front view.
Download figure:
Standard image High-resolution imageFurther, sensor configuration is such that each foot has the force sensor, and the trunk has a torque and an angle sensor. The force sensors detect normal forces Ni, and the torque sensor detects the generated torque of the rotary actuator at the trunk. An angle sensor at the trunk detects the trunk-joint angle  .
.  is negative when the right body side is concave, as shown in figure 2(b).
is negative when the right body side is concave, as shown in figure 2(b).
2.2. Control system
This section describes the decentralized control based on the feedback structure as shown in figure 1(a); the effect of sensory feedback mechanisms and their contribution to body–limb coordination is discussed.
2.2.1. Leg control
The phase of the oscillator implemented in each leg is described as follows:
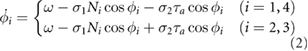
where  (rad s−1) denotes the intrinsic angular velocity of the phase oscillators,
(rad s−1) denotes the intrinsic angular velocity of the phase oscillators,  (rad (N · s)−1) and
(rad (N · s)−1) and  (rad (N · m · s)−1) are the weights of the sensory feedback terms. Ni represents the normal force detected at tip of the foot.
(rad (N · m · s)−1) are the weights of the sensory feedback terms. Ni represents the normal force detected at tip of the foot.  (N · m) represents the torque generated at the trunk actuator.
(N · m) represents the torque generated at the trunk actuator.
The second term works according to the first rule in the control scheme as shown in figure 1(a). The local feedback rule was proposed by Owaki et al [11] and generates adaptive interlimb coordination in response to speed and the physical properties of the robot [11, 12]. Based on the effect of the sensory feedback, the oscillator phase is modulated toward  when Ni > 0. This means that the leg remains on the ground when it supports the body. The local sensory information Ni describes how much the other legs support the body, such that this term can generate adaptive interlimb coordination without information from the other legs.
when Ni > 0. This means that the leg remains on the ground when it supports the body. The local sensory information Ni describes how much the other legs support the body, such that this term can generate adaptive interlimb coordination without information from the other legs.
The third term works according to the second rule in the control scheme as shown in figure 1(a). Based on the effect of the sensory feedback as shown in figure 3(a), when the trunk actuator makes the left side of the body concave ( ), the oscillator phase of the left fore and right hind legs are modulated toward
), the oscillator phase of the left fore and right hind legs are modulated toward  , and the phase of the left hind and right fore legs are modulated toward
, and the phase of the left hind and right fore legs are modulated toward  . This means that leg motions are modulated to exploit lateral bending.
. This means that leg motions are modulated to exploit lateral bending.
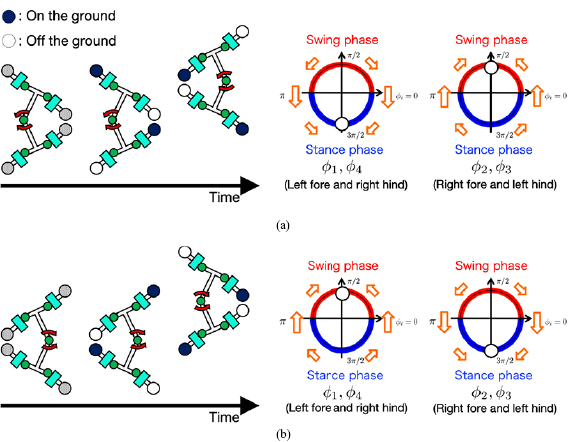
Figure 3. Sensory feedback from body to limb that modulates leg motions to exploit lateral bending. It works according to the second rule as shown in figure 1(a). (a) Trunk actuator makes the left side of the body concave ( ). (b) Trunk actuator makes the right side of the body concave (
). (b) Trunk actuator makes the right side of the body concave ( ).
).
Download figure:
Standard image High-resolution image2.2.2. Body control
The torque in the trunk joint is described as follows:
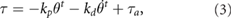
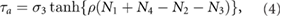
where the first and second terms originate from the passive spring damper and the third term is attributed to the rotary actuator. kp and kd denote the spring and damper coefficient.  represents the torque at the trunk actuator that operates depending on the normal forces detected by the limbs.
represents the torque at the trunk actuator that operates depending on the normal forces detected by the limbs.  (N · m) and
(N · m) and  (1/N) represent the weight of the sensory feedback.
(1/N) represent the weight of the sensory feedback.
The first and second terms are required to maintain the neutral angle of the trunk and the third term  operates as the third rule in the control scheme as shown in figure 1(a). As shown in figure 4, the trunk bends in response to ground contact. When left fore and/or right hind limbs are on the ground (
operates as the third rule in the control scheme as shown in figure 1(a). As shown in figure 4, the trunk bends in response to ground contact. When left fore and/or right hind limbs are on the ground ( ), the trunk actuator makes the left side of the body concave (
), the trunk actuator makes the left side of the body concave ( ). Similarly, when the left hind and/or right fore limbs are on the ground (
). Similarly, when the left hind and/or right fore limbs are on the ground ( ), the actuator makes the right side of the body concave (
), the actuator makes the right side of the body concave ( ). The interaction of the second and third rules in figure 1(a) establishes the relationship between the legs and the trunk, which facilitates longer strides and more powerful kicking against the ground.
). The interaction of the second and third rules in figure 1(a) establishes the relationship between the legs and the trunk, which facilitates longer strides and more powerful kicking against the ground.
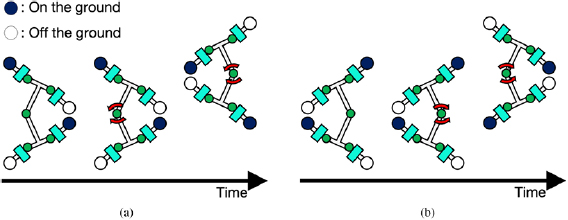
Figure 4. Sensory feedback from limb to body that actuates trunk joint to extend stride and enhance thrust. It works according to the third rule as shown in figure 1(a). (a) Left fore and/or right hind limbs are on the ground ( ). (b) Left hind and/or right fore limbs are on the ground (
). (b) Left hind and/or right fore limbs are on the ground ( ).
).
Download figure:
Standard image High-resolution image3. Simulation
3.1. Steady locomotion
We conducted robot simulations using open dynamics engine [13] and all trials lasted 60 s. In the beginning, we observed the manner of locomotion. Parameter values were determined heuristically as listed in table 1. Figures 5 and 6(a), and movie 1a (stacks.iop.org/BB/14/066010/mmedia) are snapshots, the gait patterns, and movie during resulting locomotion, respectively. The upper and lower graphs in figure 6(a) show the time evolution of the trunk-joint angle  and the gait diagrams, where the colored region represents the stance phases (Ni > 0). The proposed model exhibited stable lateral sequence walking gait with lateral bending and the expected relationship between the legs and the trunk, shown in figures 3 and 4, in few seconds without any predefined relationship. These features and the obtained gait diagram are qualitatively similar to those of sprawling locomotion shown in salamanders and lizards [3, 4].
and the gait diagrams, where the colored region represents the stance phases (Ni > 0). The proposed model exhibited stable lateral sequence walking gait with lateral bending and the expected relationship between the legs and the trunk, shown in figures 3 and 4, in few seconds without any predefined relationship. These features and the obtained gait diagram are qualitatively similar to those of sprawling locomotion shown in salamanders and lizards [3, 4].

Figure 5. Snapshots of the simulated robot model during locomotion. It bends the trunk to increase the stride length such that the model builds an expected relationship between the legs and the trunk as shown in figures 3 and 4.
Download figure:
Standard image High-resolution image
Figure 6. Time evolutions of the trunk-joint angle  (upper) and the gait diagrams (lower), where the colored region represents the stance phases (Ni > 0). (a) proposed model whose parameters are listed in table 1, mean speed is 0.099 (m s−1), (b) open loop control model predefined the relationship between the legs and the trunk without any sensory feedback and the parameters are listed in table 1, mean speed is 0.109 (m s−1), (c) rigid spine model whose parameters are the same as that of the proposed model except for the compliance of trunk-joint and the mutual feedback gain (
(upper) and the gait diagrams (lower), where the colored region represents the stance phases (Ni > 0). (a) proposed model whose parameters are listed in table 1, mean speed is 0.099 (m s−1), (b) open loop control model predefined the relationship between the legs and the trunk without any sensory feedback and the parameters are listed in table 1, mean speed is 0.109 (m s−1), (c) rigid spine model whose parameters are the same as that of the proposed model except for the compliance of trunk-joint and the mutual feedback gain ( ,
,  ), mean speed is 0.038 (m s−1), (d) passive spine model whose parameters are the same as the proposed model except for the mutual feedback gain (
), mean speed is 0.038 (m s−1), (d) passive spine model whose parameters are the same as the proposed model except for the mutual feedback gain ( ), mean speed is 0.016 (m s−1).
), mean speed is 0.016 (m s−1).
Download figure:
Standard image High-resolution imageTable 1. Parameter values of employed in the simulations.
| Parameter | Value | Unit |
|---|---|---|
| Trunk length | 0.24 | (m) |
| Shoulder/hip length | 0.13 | (m) |
| Leg length | 0.10 | (m) |
| Total mass (m) | 1.50 | (kg) |
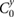 |
0.00 | (rad) |
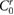 |
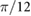 |
(rad) |
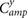 |
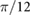 |
(rad) |
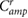 |
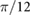 |
(rad) |
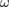 |
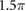 |
(rad s−1) |
 |
0.90 | (rad (N · s)−1) |
 |
2.00 | (rad (N · m · s)−1) |
 |
1.70 | (N · m) |
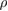 |
0.05 | (1/N) |
| kp | 1.50 | (N · m rad−1) |
| kd | 0.02 | (N · m · s rad−1) |
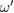 |
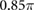 |
(rad s−1) |
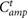 |
0.5 | (rad) |
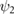 |
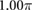 |
(rad) |
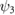 |
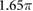 |
(rad) |
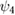 |
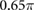 |
(rad) |
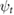 |
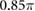 |
(rad) |
We compared the locomotion of the proposed model with those of other three models—open loop control model, rigid spine model, and passive spine model. Open loop control model predefined the relationship between the legs and the trunk and without any sensory feedback. Each leg and trunk has a phase oscillator that defines the target angle of each actuator as follows:
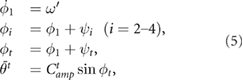
where  (rad s−1) denotes the intrinsic angular velocity of the phase oscillators,
(rad s−1) denotes the intrinsic angular velocity of the phase oscillators,  (rad) is the phase difference between the oscillators in the leg and the left fore leg.
(rad) is the phase difference between the oscillators in the leg and the left fore leg.  (rad) is the phase of oscillator in trunk and
(rad) is the phase of oscillator in trunk and  is the phase difference between the oscillators in the trunk and left fore leg. The target angles of leg actuators
is the phase difference between the oscillators in the trunk and left fore leg. The target angles of leg actuators  ,
,  are calculated using equation (1).
are calculated using equation (1).  represents the amplitude of the trunk actuator. The parameters
represents the amplitude of the trunk actuator. The parameters  and
and  was determined referring to the gait cycle and the trunk-joint angle of proposed model (figure 6(a)), and we selected the combination of phase differences
was determined referring to the gait cycle and the trunk-joint angle of proposed model (figure 6(a)), and we selected the combination of phase differences  and
and  that exhibited the highest locomotion speed of any combinations of the phase differences (
that exhibited the highest locomotion speed of any combinations of the phase differences ( ) as summarized in table 1.
) as summarized in table 1.
Figure 6(b) and movie 1b show the gait patterns that are in agreement with those in the proposed model. Although the wave shape of the time evolution of trunk-joint angle is different, the foot fall patterns, relationship between the legs and the trunk are almost same (figures 6(a) and (b)).
Rigid spine model is same as the CPG model proposed by Owaki [11, 12] that has the rigid trunk ( ) and the control algorithm without the second and third rules for body–limb coordination (
) and the control algorithm without the second and third rules for body–limb coordination ( ). It shows that the walking-trot gait and the trunk remain straight during locomotion (figure 6(c), movie 1c), which is the similar behavior observed in the previous study [12].
). It shows that the walking-trot gait and the trunk remain straight during locomotion (figure 6(c), movie 1c), which is the similar behavior observed in the previous study [12].
Passive spine model is the same as the rigid spine model except for the trunk that is flexible ( ). It shows that an inefficient locomotion in which the right fore leg placed on the ground, and the trunk bends to form the right (left) side of the body concave when the left fore (hind) leg is on the ground (figure 6(d), movie 1d).
). It shows that an inefficient locomotion in which the right fore leg placed on the ground, and the trunk bends to form the right (left) side of the body concave when the left fore (hind) leg is on the ground (figure 6(d), movie 1d).
Next, we measured the locomotion mean speed  and CoT. They are calculated as follows:
and CoT. They are calculated as follows:
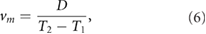
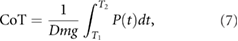
where T1 (s) and T2 (s) are the simulation times, and D (m) is distance travelled from T1 to T2. T1 = 20, T2 = 60 are defined such that sufficient time is allowed to converge a steady state. m (kg) is the total mass of the model, g (m s−2) is the gravitational acceleration, and  (W) is the power consumption of the actuators that was estimated based on the model proposed by Nishii [14]:
(W) is the power consumption of the actuators that was estimated based on the model proposed by Nishii [14]:
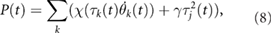
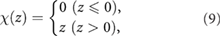
where  is a positive constant related to energy consumption owing to heat emission. We determined
is a positive constant related to energy consumption owing to heat emission. We determined  , which refers to the power consumption of the servomotor, Dynamixel XM430-W350 (ROBOTIS). We conducted 64 trials for various combinations of the initial phases of the oscillators as
, which refers to the power consumption of the servomotor, Dynamixel XM430-W350 (ROBOTIS). We conducted 64 trials for various combinations of the initial phases of the oscillators as  and
and  on each control rule. Figure 7 shows the mean speed and the CoT for each control rule, where the height of the plots and the error bars represent the mean values and standard deviations of all the trials, respectively. Proposed model achieved similar performances of open loop control model, which is a higher speed (
on each control rule. Figure 7 shows the mean speed and the CoT for each control rule, where the height of the plots and the error bars represent the mean values and standard deviations of all the trials, respectively. Proposed model achieved similar performances of open loop control model, which is a higher speed ( ) and energy efficiency (CoT <4.0) than those of rigid and passive spine models.
) and energy efficiency (CoT <4.0) than those of rigid and passive spine models.
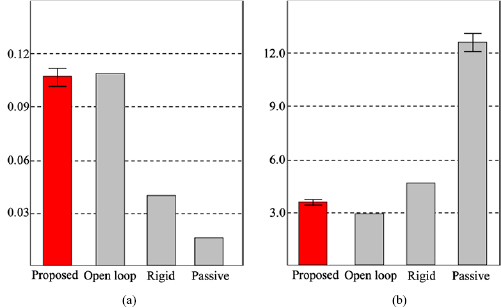
Figure 7. Locomotion performances, (a) locomotion speed of proposed model ( , SD =0.0052), open loop control model (
, SD =0.0052), open loop control model ( , SD
, SD  ), rigid spine model (
), rigid spine model ( , SD =0.0003), and passive spine model (
, SD =0.0003), and passive spine model ( , SD =0.0005), (b) CoT of proposed model (CoT =3.57, SD =0.167, open loop control model (CoT =2.89, SD =0.025), rigid spine model (CoT =4.65, SD =0.057), passive spine model (CoT =12.60, SD =0.527). Each performance is measured for each model using 64 trials for various combinations of the initial phases of the oscillators as
, SD =0.0005), (b) CoT of proposed model (CoT =3.57, SD =0.167, open loop control model (CoT =2.89, SD =0.025), rigid spine model (CoT =4.65, SD =0.057), passive spine model (CoT =12.60, SD =0.527). Each performance is measured for each model using 64 trials for various combinations of the initial phases of the oscillators as  and
and  .
.
Download figure:
Standard image High-resolution image3.2. Fault tolerance
To investigate fault tolerance, we assumed leg paralysis for our experiment. In this case, leg paralysis means that the phase of the oscillator is fixed to a certain value ( ). Figure 8 shows that gait diagrams under fore legs (
). Figure 8 shows that gait diagrams under fore legs ( ) and hind legs paralysis conditions (
) and hind legs paralysis conditions ( ), respectively. Movie 2a and 2b are movies demonstrating fore legs and hind legs paralysis conditions, respectively. Even if the positions of the fore/hind legs are fixed, the model exhibited stable walking that is a qualitatively similar gait to that under intact conditions (figure 6(a)). Figure 9 shows that locomotion speed and CoT under fore/hind legs paralysis condition in the proposed and open loop control models, where the height of the plots represent the mean values of 16 trials based on various combinations of the initial phases of the oscillators as
), respectively. Movie 2a and 2b are movies demonstrating fore legs and hind legs paralysis conditions, respectively. Even if the positions of the fore/hind legs are fixed, the model exhibited stable walking that is a qualitatively similar gait to that under intact conditions (figure 6(a)). Figure 9 shows that locomotion speed and CoT under fore/hind legs paralysis condition in the proposed and open loop control models, where the height of the plots represent the mean values of 16 trials based on various combinations of the initial phases of the oscillators as  ,
,  or
or  ,
,  . Movies 3a and 3b capture gait under fore legs and hind legs paralysis conditions in open loop control model, respectively. In the proposed model, under fore legs paralysis condition, the speed reduced by about 19% and CoT decreased by about 17.2%; further, under hind legs paralysis condition, the speed reduced by about 14.5% and CoT increased by about 42.9% that when under intact condition. In the open loop control model, under fore legs paralysis condition, the speed reduced by about 12.3% and CoT decreased by about 24.3%, and under hind legs paralysis condition, the speed reduced by about 42.0% and CoT increased by about 161.8% than when under intact condition. The result indicates that the hind leg paralysis is critical and leads to mobility failure. Despite the fatal failure, the proposed model works without losing the mobility, relatively.
. Movies 3a and 3b capture gait under fore legs and hind legs paralysis conditions in open loop control model, respectively. In the proposed model, under fore legs paralysis condition, the speed reduced by about 19% and CoT decreased by about 17.2%; further, under hind legs paralysis condition, the speed reduced by about 14.5% and CoT increased by about 42.9% that when under intact condition. In the open loop control model, under fore legs paralysis condition, the speed reduced by about 12.3% and CoT decreased by about 24.3%, and under hind legs paralysis condition, the speed reduced by about 42.0% and CoT increased by about 161.8% than when under intact condition. The result indicates that the hind leg paralysis is critical and leads to mobility failure. Despite the fatal failure, the proposed model works without losing the mobility, relatively.
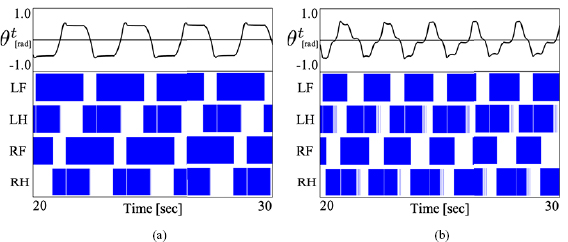
Figure 8. Gait diagrams under leg paralysis condition. Paralysis implies that the phase of oscillator is fixed at a certain value ( ). (a) paralyzed fore limbs imply that
). (a) paralyzed fore limbs imply that  , (b) paralyzed hind limbs implies that
, (b) paralyzed hind limbs implies that  .
.
Download figure:
Standard image High-resolution image
Figure 9. Locomotion performances, (a) locomotion speed, of proposed model under fore legs paralysis condition ( , SD =0.0004) and hind legs paralysis condition (
, SD =0.0004) and hind legs paralysis condition ( SD =0.0006), and of open loop control model, under fore legs paralysis condition (
SD =0.0006), and of open loop control model, under fore legs paralysis condition ( , SD
, SD  ) and hind legs paralysis condition (
) and hind legs paralysis condition ( , SD
, SD  ), (b) CoT, of proposed model under fore legs paralysis condition (CoT =2.97, SD =0.009) and hind legs paralysis condition (CoT =5.11, SD =0.034), and of open loop control model, under fore legs paralysis condition (CoT =2.19, SD =0.0003) and hind legs paralysis condition (CoT =7.58, SD =0.0022). The performance of each model using 16 trials based on various combinations of the initial phases of the oscillators as
), (b) CoT, of proposed model under fore legs paralysis condition (CoT =2.97, SD =0.009) and hind legs paralysis condition (CoT =5.11, SD =0.034), and of open loop control model, under fore legs paralysis condition (CoT =2.19, SD =0.0003) and hind legs paralysis condition (CoT =7.58, SD =0.0022). The performance of each model using 16 trials based on various combinations of the initial phases of the oscillators as  ,
,  ,
,  ,
,  ,
,  or
or  ,
,  ,
,  ,
,  ,
,  .
.
Download figure:
Standard image High-resolution image3.3. Effects of the bidirectional sensory feedback between limbs and body
A grid search with various combinations of  and
and  , which are the sensory feedback gains for body to limbs and limbs to body, respectively, was conducted to investigate the effects of feedback on the locomotion speed and CoT. Other parameters are determined as listed in table 1. Figure 10 shows the graph where a brighter colored region indicates better performance. For higher speed,
, which are the sensory feedback gains for body to limbs and limbs to body, respectively, was conducted to investigate the effects of feedback on the locomotion speed and CoT. Other parameters are determined as listed in table 1. Figure 10 shows the graph where a brighter colored region indicates better performance. For higher speed,  above a certain value is required; further,
above a certain value is required; further,  is not very influential (figure 10(a)). However, for high energy efficiency, both feedback gains are influential figure 10(b). In particular, the best performance is represented by the parameter area using both feedback rules, whereas the parameter areas that use either feedback or no feedback, e.g.
is not very influential (figure 10(a)). However, for high energy efficiency, both feedback gains are influential figure 10(b). In particular, the best performance is represented by the parameter area using both feedback rules, whereas the parameter areas that use either feedback or no feedback, e.g.  ,
,  , or
, or  ,
,  (the same as the passive spine model) tends to lower the performance. In addition, the results demonstrate that the model generates effective locomotion in a large control parameter area about
(the same as the passive spine model) tends to lower the performance. In addition, the results demonstrate that the model generates effective locomotion in a large control parameter area about  and
and  . It indicates that the proposed model has the insensibility of the control parameter toward the performance and the cross-coupled sensory feedback facilitates parameter tuning.
. It indicates that the proposed model has the insensibility of the control parameter toward the performance and the cross-coupled sensory feedback facilitates parameter tuning.
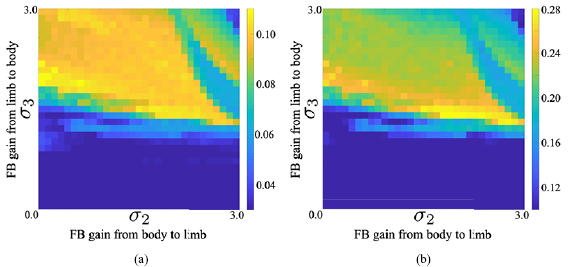
Figure 10. Effects of the bidirectional sensory feedback between the limbs and the body in terms of speed and energy efficiency. Energy efficiency is the reciprocal of CoT calculated using equation (7). The horizontal axis denotes the result of unidirectional feedback from body to limb ( ,
,  ) and the vertical axis denotes the result of unidirectional feedback from limb to body (
) and the vertical axis denotes the result of unidirectional feedback from limb to body ( ,
,  ). (a) Locomtion speed (m s−1). (b) Energy efficiency.
). (a) Locomtion speed (m s−1). (b) Energy efficiency.
Download figure:
Standard image High-resolution image3.4. Robustness to change of body structure
We observed a manner of locomotion when body structure is changed to investigate robustness to variation in body property (figure 11). The proposed model shows stable locomotion with both of elongated and shortened body without any change to the control parameters. The result indicates that the proposed model facilitates stable locomotion without precise body design.
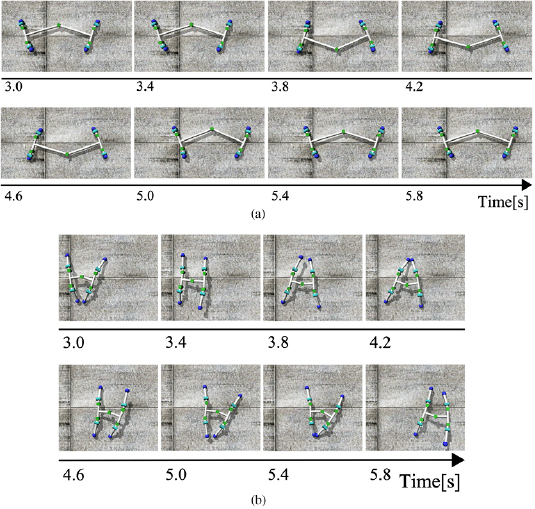
Figure 11. Snapshots of the simulated robot model during locomotion. Control parameters are listed in table 1. (a) Elongated body, trunk length 0.6 m, shoulder length 0.13 m, leg length 0.03 m. (b) Shortened body, trunk length 0.2 m, shoulder length 0.13 m, leg length 0.12 m.
Download figure:
Standard image High-resolution image4. Discussion
In this section, we discuss four issues: the usefulness of sensory feedback for body–limb coordination, effect of lateral bending in sprawling locomotion, advantage of bidirectionality in cross-coupled feedback, and application to a real salamander-like robot and other animal-like robots that employ body–limb coordination.
We initially compared the results obtained for the proposed and open loop control models to discuss the usefulness of sensory feedback for body–limb coordination. Cross-coupled sensory feedback can generate almost the same body–limb coordination as the open loop control model, which optimizes the relationship between the legs and the trunk; further, it can achieve higher speed and energy efficiency (figures 6 and 7). Furthermore, the proposed model is superior than open loop control model in terms of two factors: fault tolerance and parameter tuning. The hind limbs of quadrupeds contribute to propulsion in legged locomotion [3], and therefore, the failure of hind legs could affect the locomotion performance. Indeed, figure 9 shows that the failure of hind legs is more influential to the speed and energy efficiency than the failure of fore legs. Toward the failure, the proposed model exhibits higher endurance than the open loop model. It is inferred that the lateral bending in response to ground contact represents the kicking motion of hind legs. The result supported the use of sensory feedback in terms of the adaptability toward hind leg failure. However, the performance of the proposed model is affected under foreleg paralysis in the experiment and the reason is not apparent. A more precise investigation is required to better understand this behavior. Further, as shown in figures 10 and 11, the proposed model works when control parameters or body structure is changed. This indicates that the model has robustness to variation in control parameters and body structure, and it need not calculate the optimized relationship between the leg and the trunk. These results suggest that the sensory feedback could be a powerful tool to generate effective locomotion with body–limb coordination.
Second, we compared the results obtained for the proposed and passive spine models (figure 7) to discuss the effect of lateral bending. Although both models have a flexible spine, the results are entirely different, and the reason is because of the relationship between the legs and the trunk. The relationship in the passive spine model emerges as follows (figure 12): the left fore and/or right hind legs kick against the ground to propel the body; as a result, the leg experiences a reaction force that causes the right side of the body to form a concave shape and the bending impedes the propulsion. The impedance of propulsion due to the bending causes the passive spine model to exhibit an inferior performance compared with that of the rigid spine model. However, the mutual feedback rule tends to result in the relationship shown in figures 3 and 4. For such body–limb coordination, lateral bending supports the improvement of the stride length resulting in higher speed and energy efficiency. This coordination improving the mobility is observed in previous studies [6, 8]. This implies that the active use of trunk supporting propulsion is effective in sprawling locomotion and is consistent with biological reports [7, 15] in which hypaxial muscles have a dual role in resisting torsional forces from the legs and bending the trunk to increase the stride length as in salamander locomotion.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 12. Relationship between leg and trunk motion in a passive spine model. The lateral bending that impedes legged locomotion emerges as follows: left fore and/or right hind leg kicks against the ground to propel the body (left figure); as a result, the leg experiences a reaction force that makes the right side of the body concave (middle figure), and the bending acts to impede propulsion (right figure).
Download figure:
Standard image High-resolution image{kind=link}
Third, we discuss the advantages of the cross-coupling of sensory feedback. The previous study that proposed a spine–limb coordination controller using sensory feedback [10] demonstrated that the feedback from limb to body is useful for the spine–limb coordination. However, how sensory feedback from body to limb or the combination of the two types of feedback affects body–limb coordination remains unclear. To investigate them, we conducted an experiment, as shown in figure 10. This shows that bidirectional sensory feedback results in a more effective locomotion than unidirectional feedback from limb to body or from body to limb or than those models without feedback. Moreover, cross-coupled sensory feedback control has a large control parameter area in which the model generates effective locomotion in terms of speed and energy efficiency. The results indicate that the sensory feedback facilitates body–limb coordination to improve mobility. This may suggest that real animals also use similar sensor feedback to improve mobility.
In this article, we adopted a minimal structure for sprawling locomotion to highlight the effects of sensory feedback in body–limb coordination. The coordination based on sensory feedback exhibits mobility similar to that of a centrally wired body–limb coordination model; further, this coordination is superior than when tuning parameters and when the hind legs fail. However, the simple body structure is not suited to compare the behavior exhibited with those of real salamanders and lizards quantitatively. Therefore, we will develop a real salamander-like model and conduct a real robot experiment to compare the gait diagram and kinematics during locomotion with real animals quantitatively. Moreover, we will also explore the other type of body–limb coordination, such as leg and head or leg and tail. By exploring them, we will attempt to extract a universal principle for body–limb coordination in quadrupeds.
5. Conclusion
This study aimed to better understand the underlying mechanism of body–limb coordination in sprawling locomotion that is exhibited on ground by salamanders and lizards using a synthetic approach. In particular, we investigated the role of sensory feedback in body–limb coordination and the extent to which sensory feedback improves the traveling ability. The proposed model has a cross-coupled sensory feedback that leads to proper leg–trunk coordination without a predefined relationship. The gait is qualitatively similar to sprawling locomotion and reaches almost the same locomotion speed and energy efficiency of a neural network model optimizing the relationship between the legs and the trunk. Moreover, cross-coupled sensory feedback helps adapt to leg failure and facilitates parameter tuning. These results suggest that sensory feedback mechanisms facilitate sprawling locomotion and that cross-coupled sensory feedbacks play an important role in body–limb coordination. Our future works will develop a real salamander-like model for quantitative comparison with the gait of salamander locomotion. In addition, we will examine the behavior of other animals in an attempt to establish a universal principle with respect to agile animal locomotion with body–limb coordination.
Acknowledgment
This work was supported by the Human Frontier Science Program (RGP0027/2017), Japan Science and Technology Agency, CREST (JP-MJCR14D5) and the Division for Interdisciplinary Advanced Research and Education, Tohoku University. The authors would like to thank Dr Akira Fukuhara and Mr Kotaro Yasui, Tohoku University, for their helpful suggestions.