
Abstract
Chimney swifts (Chaetura pelagica) are highly aerial, small, insectivorous birds well known for roosting en masse in chimneys during their autumn migration. These roosting events require hundreds to thousands of birds to enter a small opening (here 0.64 m2) within a short amount of time (15–30 min). Thus, these entry events pose a complex navigational and behavioral challenge as birds identify their entry route, cooperate with other birds present to form an entry flock, and compete with other birds at the time of chimney entry. We used six synchronized cameras to capture and reconstruct the 3D flight trajectories of swifts before and during chimney entry. Navigation into the chimney is consistent with use of a relative retinal expansion velocity cue, which results in an entry/non-entry decision point about 1.5 m above the chimney, or 0.4 s at typical entry speeds. Entries were highly clustered with 91 of 136 entries occurring within 1 s of another entry. We observed both synchronous (entry within 0.2 s) and sequential entry behavior (entry separated by ~0.4 s). Birds entering the chimney flew in close proximity to other birds (median minimum distance 0.51 m; 1.7 wingspans). In cases where two birds appeared to attempt a near-simultaneous entry, the bird either slightly to the rear or with a velocity vector bringing it closer to the current position of the other bird tended to make an avoidance maneuver and abandon its entry attempt. Overall, these results show how groups of animals execute complex landing and collision avoidance maneuvers in a natural setting without central control authority.
Export citation and abstract BibTeX RIS
View the Video Abstract for this article.
Introduction
Chimney swifts are notable for their overnight roosting in chimneys alongside hundreds to thousands of fellow swifts. Near dusk, the swifts gather in flocks that circle the chimney before making a rapid approach inside. Although the overall behavior is well known, quantitative investigation of the group behavior and flight biomechanics of this unusual bird aggregation has just begun, with a single study examining the overall behavior and characteristic biomechanics [1] for a large roosting flock of ~1000 birds. That study was not able to resolve the final entry trajectories of the swifts, but nevertheless concluded from the decline in correlation of flight trajectories as birds approached and entered the chimney that rather than cooperatively following one another into the chimney, birds competed for entry positions. Here, we examine the detailed entry trajectories of a smaller chimney swift flock with ~130 roosting birds to better understand how swifts guide themselves into the chimney, when they make a final decision to enter, whether their presumed visual guidance mechanism is affected by the decreasing light levels during the course of chimney entry, and how competition during entry manifests in the flight trajectories of entry versus non-entry outcomes of birds approaching the chimney.
Although little is known about the landing process of chimney swifts, visually guided landing behavior has been investigated in several other aerial species, from flies to hawks [2–4]. From video recordings of female houseflies (Musca domestica) landing on stationary spheres, Wagner [2] calculated several optical and metric distance parameters, including distance, retinal angle of the sphere (α) and relative retinal expansion velocities (RREV) in an effort to determine what cue controlled the onset of the landing phase (See Methods for detailed descriptions). Among the variables measured, RREV exhibited the least variation just prior to landing initiation and was therefore identified as a trigger for the maneuver. During fly landings, RREV increased monotonically, indicating that the time point of minimum coefficient of variation corresponds to a threshold value of RREV that is used for initiating landing.
RREV is an example of a tau function [4], which provides a first-order approximation of the time-to-contact (RREV equals the inverse of tau, the time to landing). RREV has also been supported as relevant to the landing behavior of hawks [3]. A recent re-investigation of landing behavior in flies [5] provided renewed support for use of RREV as a cue for insect landing maneuvers [5], while also noting the difficulty of differentiating RREV specifically from similar landing models in observational data. Based on these data, we hypothesize that swifts use RREV as a cue for coordinating chimney entry, and that this RREV threshold would also be used for deciding whether to enter the chimney. In principle, RREV is independent of flight speed and approach height, so even if these quantities vary through the evening (possibly due to changes in lighting), we do not expect any characteristic RREV to change, even though metric distances associated with it might vary.
Quantitative examination of a different and larger chimney swift entry flock revealed that chimney entries occurred almost continuously once birds began landing, and that near-simultaneous entries came from birds on divergent flight trajectories, making the flock cooperative but actual chimney entry competitive [1]. Following these results, we hypothesized that the smaller flock would also exhibit competitive entry, with near simultaneous entries occurring from different directions if they occurred at all.
Finally, the earlier quantitative flocking study showed that chimney swifts maintain an avoidance zone of approximately 1.4 m, equivalent to ~4.5 of their 0.3 m wingspans. It is unclear whether swifts maintain this avoidance zone during chimney entry, which is likely to bring birds close to one another as their trajectories converge. Mass chimney entry likely requires simultaneous mitigation of collision risk and maintenance of a trajectory toward the chimney. Cliff swallows (Petrochelidon pyrrhonota), another avian aerial insectivore, have been shown to use avoidance trajectories consistent with a collision cone model [6], but it is unclear how such a model will arbitrate between the dual problem of collision avoidance and chimney entry. In cases where a possible collision requires a large-magnitude avoidance maneuver, it is unknown what (if any) factors give one bird precedence over the other, or whether both birds react similarly. A precedence rubric should allow one of the birds to continue toward collision entry, whereas purely reactive collision avoidance is likely to lead to neither bird making an entry. Based on previous collision cone results [6] and earlier research on swallows showing that birds in both front and rear positions can influence one another's trajectories [7], we predicted that no collision avoidance rubric exists and that close encounters near the chimney would result in neither bird landing.
Answering these questions reveals how birds cooperate and compete to execute the impressive feat of group entry into a small (0.64 m2) landing with a median time of 0.57 s between entries. Interest in biomimetic landing strategies for micro-air vehicles (MAVs) has largely been confined to replication of perching behavior [8], but the increasing emphasis on MAV swarms [9, 10] suggests that an understanding of the guidance mechanisms underlying massed landings by flying animals, especially landings in a confined space such as chimneys or caves, may also serve to inspire new strategies for MAV swarm control.
Methods
Videography
We recorded continuous footage of chimney swifts flying around and entering a chimney in Raleigh, NC on September 20, 2014 using three Canon EOS 6D cameras (1920 × 1080 pixel resolution; 29.97 frames per second) and three GoPro Hero3 Black Edition cameras (1280 × 720 pixel resolution; 119.88 frames per second). Canon cameras were placed 20 m from the chimney and with 9.4 m between the outermost cameras to provide a broad lateral view of the scene (figure 1). GoPro cameras were placed on three sides of the chimney looking up to provide a detailed view of the birds as they entered the chimney (figure 1). All six cameras were synchronized using audio tones that were broadcast using portable radios (Motorola MH230R) [11]. Birds were not handled or interfered with during recording, and the observation protocol was approved by the University of North Carolina Institutional Animal Care and Use Committee.
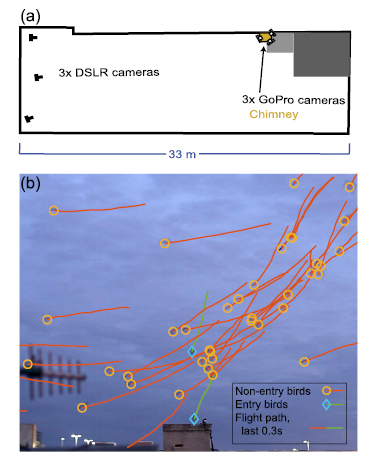
Figure 1. Recording setup and example videos (a) shows an overhead view of the recording setup, with 3 Canon DSLR cameras providing a lateral view and 3 GoPro cameras providing a vertical view of chimney entry. (b) Shows a sample frame from a DSLR, cropped to close to the chimney and showing the position and trajectory of non-entry and entry birds. See supplemental movie 1 for an animation of (b).
Download figure:
Standard image High-resolution image3D calibration and reconstruction of flight paths
The cameras were calibrated using a 0.94 m wand passed through a portion of the recording volume near the chimney [12]; additional shared view information was provided by the manually tracked trajectories [13] of seven birds that flew through the recording volume from different directions, thus extending the calibrated volume to include the space used by the birds and encompassing an approximately 58 000 m3 volume. The joint six camera calibration was centered on the chimney and was aligned to gravity using the chimney edge, visible in the three Canon cameras (figure 1). The coefficient of variation of wand length (i.e. the standard deviation of length divided by mean length) was 1.8%. Flight paths were reconstructed automatically from the three broad lateral view cameras using methods similar to [1]. Briefly, the workflow first identifies moving foreground points in each image (i.e. the birds) as regions of large magnitude optic flow, then associating these detections among cameras by finding sets of three detections that satisfied a minimum reprojection error threshold when used to compute a 3D point. These 3D points were then associated through time using Hungarian assignment minimizing the overall difference between observations in frame n and the predicted location of birds tracked in frame n − 1. Kinematics from automatic reconstruction were sampled at 29.97 Hz to make use of the wider viewpoint of the Canon cameras (figure 1).
Data analysis
For each bird track we calculated the distance from the center of the chimney opening (d), chimney closing speed (s), total flight speed, horizontal flight speed, vertical flight speed, and the chimney approach angle (ϕ), which is the 3D angle between the bird's current trajectory and the vector to the chimney center. We also calculated variables related to visual information that the swifts could potentially use to coordinate their landing [2, 5]. The retinal size of the chimney (α) is the angle subtended by the chimney on the bird's retina, which is calculated as:
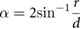
where r is half the width of the chimney. The retinal expansion velocity, Ω, is:
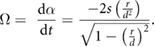
We tested the hypothesis that swifts use the relative retinal expansion velocity as a cue for coordinating their entry into the chimney. RREV is calculated as:
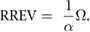
For comparison of successive entry trajectories, we used the difference in 3D entry trajectories (dϕ) between pairs of birds. We computed (dϕ) by first fitting a 3D line to the last 15 positions before entry (0.5 s, typically 1.5– 2.0 m of flight trajectory) of each entry bird using principle components analysis (PCA). The difference in 3D entry trajectory (dϕ) for each bird is then the angle between its entry vector and the entry vector of the preceding bird. See figure S1 (stacks.iop.org/BB/14/055005/mmedia) for example trajectories and their PCA fit.
For analysis of possible collision avoidance events we calculated the minimum distance of each bird to its neighbor and neighbor angle  , the 3D angle between the focal bird's velocity vector V and the vector to its neighbor N:
, the 3D angle between the focal bird's velocity vector V and the vector to its neighbor N:
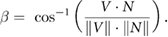
For detailed analysis of collision risk and avoidance during entry, we identified 'neighbor pairs'. These were pairs of birds that were each other's minimum distance neighbor, had a minimum bird–bird distance of <0.6 m, were observed during group entry times (i.e. within 0.5 s of a chimney entry event), had their minimum distance interaction less than 3 m and greater than 0.5 m above the chimney and had a 'close approach' to the chimney as determined by maximum RREV (see below). These stipulations served to restrict the neighbor pairs dataset to cases consistent with the birds interacting and both attempting to enter the chimney.
Results
We captured video of a 22 min period starting at 19:22 (h:m) during the evening of September 20, 2014 when swifts were flying near and then entering the chimney. This produced a total of 16 992 chimney swift flight trajectories. We observed a maximum of 159 birds present at one time. A total of 136 birds flew into the chimney and seven birds flew out of the chimney after initially entering. The first entry was observed at 19:24 and the last entry at 19:42.
Chimney approach behavior
Over the 2 s prior to entry, swifts gradually slowed flight their overall flight speed while increasing their vertical (downward) speed as they descended into the chimney (figures 2(a)–(c)). Flight speed was mostly constant for the last 0.5 s before entry. Flight speeds were highly variable between birds, ranging from 3.9 to 10.8 m s−1 at 2 s before entry and 1.6 to 7.3 m s−1 at 0.5 s before entry. Birds entering earlier in the night tended to fly faster (Linear regression; N = 135; R2 = 0.19; F = 32.4; d.f. = 134; P < 0.000 01; speed averaged over last 1 s before entry). We also examined the chimney approach angle (ϕ) as a function of time to entry (figure 2(d)). Median ϕ values were approximately 35° from 1.8 to 1.2 s prior to entry before decreasing steadily to ~8° (i.e. nearly direct approach to chimney center) 0.3 s before entry.
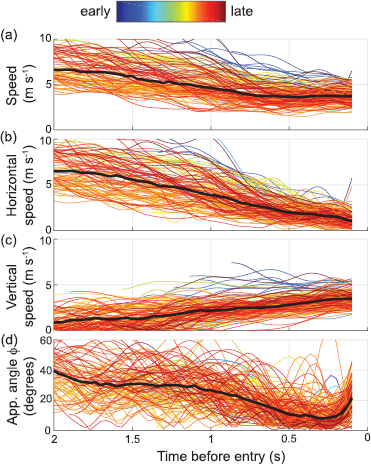
Figure 2. Characterization of entry flights. 3D (a), horizontal (b) and vertical (c) flight speed and chimney approach angle ϕ (d) for the two seconds preceding entry into the chimney from 136 trajectories. Lines are colored to indicate time of night. Thick black lines show medians. Chimney approach angle ϕ is the angle between the swift's instantaneous velocity vector and the vector from the bird to the center of the chimney opening.
Download figure:
Standard image High-resolution imageRelative retinal expansion velocity (RREV)
The coefficient of variation in RREV reached a distinct minimum value 0.4 s before entry, corresponding to an RREV value of 2.4 (figure 3). The minimum coefficient of variation in RREV (0.12) was considerably less than that of either α (0.24) or Ω (0.26), indicating that as expected, RREV provided more consistent information regarding the approach to the chimney than either α or Ω alone. The minimum coefficient of variation in RREV occurred 0.4 s prior to entry, a time that corresponds to when birds reach a roughly constant speed and low chimney approach angle (figure 2). This result supports the hypothesis that swifts use RREV, or similar visual processing cues that cannot be differentiated from RREV, for deciding when to coordinate their final chimney entry maneuver. The time of minimum coefficient of variation in RREV did not change throughout the night, when binning entries occurring within 30 s of one another (linear regression; N = 10; P = 0.65).

Figure 3. Relative retinal expansion velocity (RREV) for the 2 s preceding chimney entry. Here we show the RREV for the 136 entry trajectories in figure 2 (left axis) and their coefficient of variation (c.v., right axis, dashed line). Note the distinct minimum in the RREV c.v. at 0.4 s before entry, demarcated by the vertical dashed line. RREV lines are colored based on time of night, following the color scheme from figure 2. The solid black line indicates median values.
Download figure:
Standard image High-resolution imageApproach behavior of non-entry birds
A histogram of the maximum RREV values of all non-entry flights (12 597 trajectories) shows two distinct peaks at 0.0 and 0.8 s−1 (figure 4). We use the valley between these peaks to differentiate events where birds either did or did not approach the chimney. We subdivide the approaches as either close approaches or far approaches depending on whether the maximum RREV value exceeds the RREV value from entry flights having minimum coefficient of variation (RREV = 2.4; figure 3). The overall peak of all approaches occurring at 0.8 s−1 suggests most non-entry approach flights achieved RREV values considerably less than the putative RREV value for coordinating final entry. Maximum RREV values of non-entry flights increased over the course of the recording period (linear regression; R2 = 0.04; f = 329.9; d.f . = 12 583; P < 0.0001), indicating that birds approached progressively closer to the chimney as the night progressed.
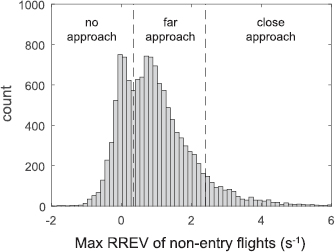
Figure 4. Histogram of maximum RREV values for birds not entering the chimney. Flights were categorized as one of three categories based on RREV values. Close approaches exceeded the RREV value with minimum coefficient of variation from entry flights (see figure 3). Far approaches had RREV below this value but above the local minimum separating the two peaks (RREV = 0.35). No approaches had RREV values <0.35. Birds that did enter the chimney had a maximum measured RREV ranging from 8.51 to 96.96.
Download figure:
Standard image High-resolution imageTiming of entries and non-entries
We examined the timing of all documented flights, far approaches, close-approaches and entries by compiling cumulative distribution functions (CDF) across the recording period (figure 5). Overall flight activity preceded all other events, followed by far approaches, close approaches and finally entries. Whereas overall flight activity and non-entry approaches were continuous from 6 to 17 min into the recording, entries occurred in bursts, as indicated by steps of the entry CDF between 12–17 min into the recording.
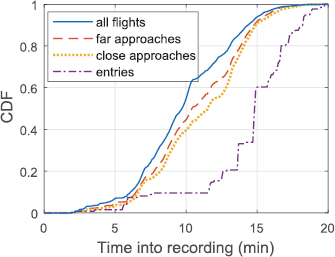
Figure 5. CDF of all flights, far approaches, close approaches, and entries during the recording period. Here we show the CDF of flights, approaches and entries during the recording period beginning at 17:22 in the evening. After an initial quiet period, flight activity is continuous throughout. Approaches begin slightly after flight activity but occur continuously thereafter. In contrast, entries occur in discrete groups visible as the rapid vertical jumps in the entries CDF.
Download figure:
Standard image High-resolution imageWe examined the timing of entries in more detail using a histogram of the time-intervals between sequential entries (figure 6). Most entries (91/136; 67%) were preceded by another bird entering the chimney within 1 s. There does not appear to be a minimum time interval between sequential entry; in 41 cases a bird entered less than 300 ms after the prior bird and in 13 cases a bird entered less than 100 ms after the prior bird. The minimum observed time between entries was 0 ms, i.e. simultaneous at the 33.4 ms granularity of our recording. The high temporal synchronization of entry behavior is noteworthy given that the 136 entries occurred over a period of 17.9 min.
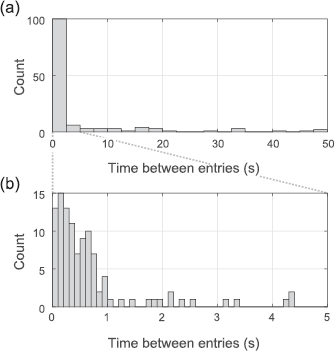
Figure 6. Histograms of the time intervals between sequential entries. Time intervals between sequential entries are shown for an expanded range of intervals (a) and for a for a narrower range (b). Note that three intervals occurred beyond the range shown in (a), with values of 99, 121 and 235 s. The median time between entries was 0.57 s.
Download figure:
Standard image High-resolution imageIn contrast to entries, only a small proportion of non-entry approaches (311/3767 far approaches, 8.3%; 154/909 close approaches, 16.9%) occurred within 1 s after an entry. This result is confirmed by visual inspection of the timing of entries and approaches in figure 5. This indicates that most non-entry approaches were not aborted because of other birds entering the chimney. Nonetheless, close approaches were more likely to be preceded by an entry within 1 s compared to far approaches (Fisher's exact test; P < 0.0001), supporting the hypothesis that some non-entry close approaches could have been aborted because of other birds interfering with their approach.
Coordination of simultaneous and sequential chimney entry
We aimed to determine how birds coordinate entry into the chimney with short time intervals separating entries. Overall, pairs of birds entering the chimney within 1 s did so by approaching from similar 3D approach angles (ϕ) (figure 7). Birds entering the chimney within 1 s did so with lower dϕ than birds entering with intervals > 1 s (figure 7(b); Mann-Whitney U test; N1 = 91; N2 = 44; U = 1302; P = 0.001). Close examination of the relationship between dϕ and time between entries for birds entering within 1 s (figure 7(a);) reveals a quadratic relationship (regression; dϕ = 20.1–37.7dt + 44.4dt2, R2 = 0.12; P = 0.004) with dϕ having a minimum value at approximately 0.4 s, and dϕ being greater closer to 0 s and 1 s. This indicates that birds entering the chimney nearly simultaneously do so by approaching from slightly different directions (see example in figures 8(a)–(c)), whereas birds exhibit the closest following behavior when entries are separated by ~0.4 s (see example in figures 8(d)–(f)). Even when birds appear to follow one another to the chimney, their exact flight paths differ slightly (figures 8(d)–(f)).
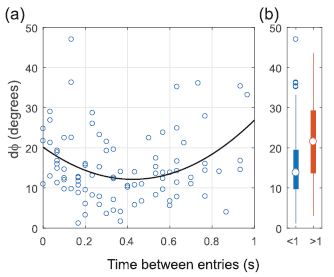
Figure 7. Difference in 3D approach angle to the chimney (dϕ) for pairs of birds entering the chimney at different time intervals. (a) For paired entries occurring with time interval dt < 1 s, dϕ was best fit as a quadratic function of time between entry (regression; dϕ = 20.1–37.7dt + 44.4dt2, R2 = 0.12; P = 0.004). dϕ was lowest for pairs of birds entering the chimney with a delay of approximately 0.4 s, indicating that these birds likely approached the chimney on similar trajectories. (b) Entries occurring within <1 s of each other had lower dϕ than entries occurring >1 s apart (Mann-Whitney U test; N1 = 91; N2 = 44; U = 1302; P = 0.001).
Download figure:
Standard image High-resolution image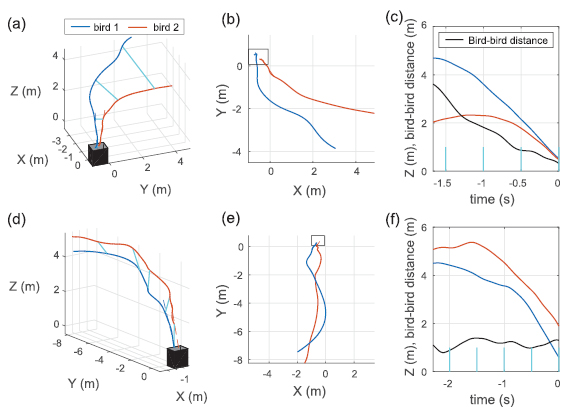
Figure 8. Example trajectories of paired-bird entries. (a)–(c) Bird 1 and bird 2 enter at the same frame and with dϕ = 21.7°. Note that the distance between birds decreases over time, with a minimum value of 0.32 m at entry. (d)–(f) Bird 1 enters 0.37 s before bird 2 and with dϕ = 4.1°. Note that the distance between birds stays relatively constant at approximately 1 m. Cyan lines connect bird trajectory positions at equivalent times and the dashed lines show the average flight path over the last 0.5 s prior to entry. See figure S1 for an enlarged 3D view.
Download figure:
Standard image High-resolution imageCollision avoidance versus entry
Birds entering the chimney had a median minimum neighbor distance of 0.51 m during approach (0.31–0.82 m quartiles). While still more than one wingspan, this value was substantially less than the previously established avoidance distance of 1.45 m for birds circling in a flock above the chimney [1] and also significantly smaller than the minimum nearest neighbor distance of birds that closely approached the chimney but did not enter (unpaired t-test, P < 0.0001).
We identified 213 'neighbor pairs' of birds that were each other's minimum distance neighbor and met other selection criteria designed to find birds that attempted chimney entry (see Methods). Of these 213 pairs, neither bird entered in 190 cases, one bird entered in 22 cases and both birds entered in 1 case. Given the total number of entries and non-entries in the neighbor pairs set, the number of single, dual and non-entries was not significantly different than random pairing. In the 22 cases where both birds may have attempted entry but only one of the pair entered the chimney, we examined each bird's neighbor angle β (the angle between its velocity vector and the vector to the other bird) relative to the time point of minimum distance between the two birds (figure 9). The average β for the bird that entered the chimney was approximately 90° throughout the interaction. In contrast, the bird that did not enter the chimney had an average β that reached a minimum of ~60° about 133 ms before its minimum distance interaction with its neighbor. β was significantly less in non-entry than entry birds at this time point (Wilcoxon rank test; P = 0.0221, n = 22) indicating that non-entry birds tended to either trail their entry partners or were flying toward them. Non-entry bird β increased rapidly starting 133 ms before the birds reached their minimum neighbor distance time and exceeded 140° as the non-entry bird flew away from the chimney and entry bird (figure 9; also see example in figure 10).
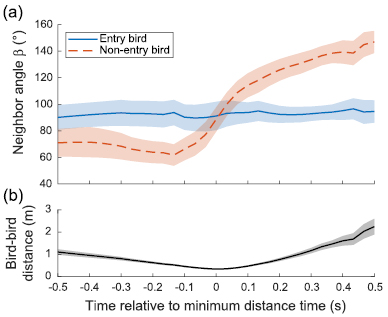
Figure 9. Neighbor angle and bird–bird distance in entry/non-entry neighbor pairs (a) shows how neighbor angle  varies with time for pairs of birds that are each other's nearest neighbor and where one bird enters but the other does not. Neighbor angle for the non-entry bird is less than the entry bird, indicating it is either behind or flying toward its neighbor. Shading indicates the standard error about each line, sample sizes range from 22 at time = 0 s to 9 at time = 0.5 s because not all tracks are of the same length. (b) shows the distance between the two birds through time.
varies with time for pairs of birds that are each other's nearest neighbor and where one bird enters but the other does not. Neighbor angle for the non-entry bird is less than the entry bird, indicating it is either behind or flying toward its neighbor. Shading indicates the standard error about each line, sample sizes range from 22 at time = 0 s to 9 at time = 0.5 s because not all tracks are of the same length. (b) shows the distance between the two birds through time.
Download figure:
Standard image High-resolution image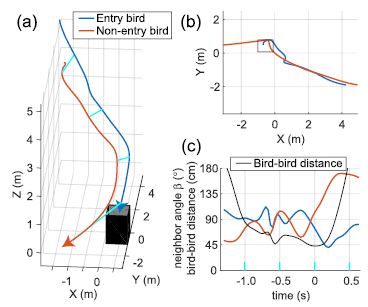
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 10. Example entry/non-entry neighbor pair interaction (a) shows the 3D trajectory of two birds which are each other's minimum distance neighbor as they approach the chimney. One bird enters and the other does not. The cyan lines connect bird positions at equivalent points in time. (b) Shows the 3D view in (a) from above; the chimney opening is shown as a rectangle. (c) Is a time series with time = 0 the time of minimum bird–bird distance. The non-entry bird has the lower neighbor angle  in prior to each of the local minimums in bird-bird distance and eventually makes a final maneuver away from the chimney.
in prior to each of the local minimums in bird-bird distance and eventually makes a final maneuver away from the chimney.
Download figure:
Standard image High-resolution image{kind=link}
Discussion
Here we quantify how more than 100 swifts coordinate entry into a 0.64 m2 chimney opening under natural conditions. We show that over the course of the landing flock formation and entry, birds use a wide range of speeds and approach trajectories, but that these approaches generally share characteristics of reducing overall speed and trading horizontal for vertical velocity during the last 2 s of approach to the chimney (figures 2(a)–(c)). We find support for the hypothesis that swifts use relative retinal expansion velocity (RREV) to coordinate their final chimney entry maneuver (figure 3), with the birds using an RREV threshold of 2.4 s−1, which corresponds to a time-to-entry (τ) of 0.4 s. The time when this occurs corresponds to the time when bird flight speed becomes constant, and chimney approach angle (ϕ) is near its minimum (figure 2(d)). We note that the RREV model is similar to other landing models such as the retinal size-dependent expansion threshold model and the time-to-contact model [5], and therefore, swifts could be using cues other than RREV for coordinating this behavior. It is not possible to differentiate among these similar visual-cue models with our data. Non-entry approaches also exhibited higher maximum RREV values as flock landing progressed, indicating that birds came progressively closer to the chimney later in the night. This might indicate that birds are learning their approach trajectories before later entering the chimney, that the decreasing light level or more crowded chimney later in the evening requires a closer approach for visual inspection. These same factors may also relate to the decrease in flight speed at entry observed in later entering birds.
We hypothesized, based on a previous analysis of chimney swift flocking, that although flock formation might be cooperative, the final chimney entry events would be competitive, e.g. birds would not follow one another into the chimney and successive entries would be from different directions. To assess these hypotheses, we documented many flights around the chimney and approaches to the chimney in addition to entries. Flight activity and non-entry approaches were nearly continuous for a 10 min period (5–15 min in figure 5) that culminated with several events from 12 to 18 min where groups of birds entered the chimney with short time delays between sequential entries (most delays between entries <1 s; figure 6).
Birds entering the chimney within 1 s approached the chimney using more similar approach angles (i.e. low dϕ), compared to birds entering with >1 s between entries (figure 7(b)). This suggests that birds entering the chimney in quick succession follow one another into the chimney, conflicting with our initial hypothesis, as does the noticeable clumping of entries in time (figure 5); birds appear to choose to enter if they observe another bird entering. The time interval where dϕ was minimized was 0.4 s (figure 7(a)). This points to an optimal time interval between two birds entering the chimney sequentially from the same direction (see example in figures 8(d)–(f)). While 0.4 s is a short time interval, at average entry speeds of 3.5 m s−1, the 0.4 time interval leaves a spacing of ~1.4 m (4.6 wingspans) between entry birds, similar to the avoidance distance of 1.45 m measured in flocking chimney swifts [1]. In contrast, birds entering the chimney nearly simultaneously (see example in figures 8(a)–(c)) had slightly higher dϕ, which appears necessary to allow co-entry without collision. This portion of the result is consistent with our initial hypothesis that successive entries would be competitive. It is also consistent with an earlier analysis of roosting flock behavior in a larger chimney swift flock [1], which found that the paths of successive entry birds diverged, i.e. had higher dϕ. In that flock, the median time between entries was ~0.2 s, an entry tempo also correlated with higher dϕ here (figure 7). These results demonstrate that swifts modify their entry behavior depending on their proximity to conspecifics.
We hypothesized that birds would avoid collisions with other birds during approach to the chimney, and that these avoidance maneuvers would tend to inhibit chimney entry. This was not the case; birds entering the chimney had a minimum nearest neighbor distance significantly less than birds that made a close approach to the chimney (defined by RREV) but did not enter. These smaller nearest neighbor distances likely enhance collision risk for entry birds. Furthermore, the birds slow down prior to entry and because aerodynamic forces are proportional to the square of airspeed, the lower flight speeds reduce the ability of the birds to generate maneuvering forces and should further increase collision risk. Finally, as a group, swifts are poorly adapted for low speed maneuvering flight, with limited control of the antebrachium [14]. Thus, chimney entry likely increases collision risk and requires that birds inhibit typical bird–bird collision avoidance responses to successfully complete the maneuver. For example, in the sequence shown in figures 8(a)–(c), the two birds gradually approach one another as they enter the chimney at approximately the same time, closing within one wingspan of one another.
However, the finding that close proximity to other birds occurs during entry does not necessarily mean that collision avoidance is not occurring or is not detrimental to entry attempts as hypothesized. To more directly address this hypothesis, we examined 'neighbor pairs' (see Methods) of birds that closely interacted and each appeared to attempt an entry. Most flights past the chimney did not result in an entry, and this was also the case for the neighbor pairs dataset. Single and paired entries were also no more or less common than expected based on the number of birds sampled. However, in cases where only one bird of the pair entered, we did find a consistent collision avoidance signal in the neighbor angle β (figure 9). Non-entry birds flew with lower neighbor angles, β, before the time of minimum bird–bird distance, indicating flight toward the neighboring bird. In contrast, entry birds maintained β of approximately 90°, keeping the neighboring bird lateral to its flight path. Non-entry birds also made a clear avoidance maneuver, whereas entry-birds did not (as indicated by no change in β for entry birds at time of minimum neighbor distance; figure 9). This suggests that a reactive collision avoidance strategy could produce the neighbor pair entry/non-entry outcome in these cases, and that the bird with the lower β is more likely to make the avoidance maneuver. However, this does not support any specific rubric, such as difference in elevation or speed, for determining which bird gets precedence when two (or more) attempt entry.
Our results show how groups of swifts execute complex landing maneuvers while avoiding collisions with conspecifics. Swifts choose to enter the chimney in close temporal and spatial proximity to conspecifics, even when there is ample time and space for birds to enter the chimney alone. This close proximity does appear to disrupt entry attempts in some cases. We propose multiple potential benefits that could offset the costs of group entry into the chimney. First, birds could use information from conspecifics to help coordinate chimney entry. Evidence of following behavior (figures 7 and 8) indicates that many birds at least partially use information from other birds' flight trajectories to coordinate their own entry into the chimney. This could be an example of collective decision-making driving movement patterns [15]. Another possibility is that sequential entry is an anti-predator behavior [16]. Nightly entries into the same chimney could put swifts at increased predation risk because predators could learn the time and location of swift entries. Following another bird into the chimney could decrease this predation risk because the predator might be occupied with trying to capture the preceding bird. Anecdotally, we have observed that swifts will abandon a chimney roost for the night if a peregrine falcon is in the vicinity, suggesting that these birds are aware of predation risk.
From a biomimetic point of view, our results further demonstrate how coordinated behavior can be produced in the absence of central control or even specific interaction rubrics for which individuals get precedence in their actions. In this case, we believe that the relative rarity of entry compared to fly-by events is one of the keys to chimney entry by the flock of swifts. Our results show that birds entering the chimney come closer to their neighbors (and presumably experience increased collision risk) than birds that merely make a close approach to the chimney but do not land. This suggests that the entry birds have temporarily suppressed their collision avoidance responses and instead rely on their neighbors maintaining these responses to prevent collisions from occurring. If several birds suppressed their avoidance responses at once a collision might ensue, so decisions to land must remain rare. However, the rarity of entries relative to close approaches means that no specific landing flight pattern is needed beyond the overall flock structure whereby birds generally fly in a similar direction and at a similar speed, a situation likely produced by avoidance of near neighbors and alignment to more distant neighbors. Our results also show that birds may share information amongst themselves during landing, e.g. birds landing in quick succession typically land along the same trajectory, but birds present at the same time already have correlated flight trajectories because of the nature of the flock so it is unclear whether directly copying or following of leader trajectories is required. Our results also show that entries are clumped in time such that one bird entering the chimney appears to increase the likelihood of another bird entering in the next few seconds. As described above, we believe this is most likely related to predator avoidance by the birds. A successful landing by one bird indicates that it is safe for other birds to try as well, and many entries in quick succession could overwhelm the capture capabilities of a solitary predator. Because engineered systems would not be subject to the same concerns, a time-varying probability for attempting entry as exhibited by the birds is likely unnecessary. Finally, because our results suggest that chimney swifts rely heavily on vision based reactive collision avoidance to enable landing, further study of the details of these responses as they are used in complex natural environments and dense groups should be informative.
Data availability
The underlying 3D flight trajectory data used to generate the results described above have been deposited in FigShare at https://doi.org/10.6084/m9.figshare.9549071.
Acknowledgments
We wish to thank Dennis Evangelista, Pranav Khandelwal and Jonathan Rader for collecting the video recordings of the chimney swift landings. The manuscript also benefitted from comments by two anonymous referees. The work was funded by the National Science Foundation, grant IOS-1253276 to TLH and by the Office of Naval Research grant N0001410109452 to TLH and others.