
Abstract
We model recent experiments on living sulphur bacteria interacting with quantised light, using the Dicke model. Our analysis shows that the strong coupling between the bacteria and the light, when both are treated quantum-mechanically, indicates that in those experiments there is entanglement between the bacteria (modelled as dipoles) and the quantised light (modelled as a single quantum harmonic oscillator). The existence of lower polariton branch due to the vacuum Rabi splitting, measured in those experiments for a range of different parameters, ensures the negativity of energy (with respect to the lowest energy of separable states), thus acting as an entanglement witness.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Witnessing quantum effects in living systems was long considered an impossible task, even by the pioneers of quantum theory, such as Bohr [1]. Recent advances in theoretical and experimental techniques, however, are bringing us closer to accomplishing it. Entanglement has extensively been investigated, and even detected, in various many-body systems [2]. Since living systems are (most probably) special cases of many-body systems, they can (presumably) be analysed with the same methods. In this paper we model recently performed experiments where living sulphur bacteria are entangled with a quantised field of light [3]. This is particularly exciting, since the quantised nature of light was at the heart of the complementarity that Bohr thought would ultimately make it impossible for us to detect quantum effects in a living entity. We also offer an argument that semi-classical models would be insufficient to explain the experiments' results.
Summary of the experiment
Let us first summarise the basics of the experiments in [3]. Green sulphur bacteria are found in anaerobic environments rich in sulphur compounds, such as microbial mats and around hot springs [4]. They are photosynthetic and are able to survive in extreme locations where light intensity drops to only a few hundred photons per second per bacteria [5]. The bacteria have evolved antenna complexes called chlorosomes, which are large aggregates of approximately 200,000 self-assembled bacteriochlorophyll (BChl) molecules. When light is absorbed by these antenna complexes, an exciton is created, which then travels to a protein baseplate attached to the chlorosome, and then to the reaction centre, where it is used to power chemical reactions.
Each bacterium contains around 200 chlorosomes, where the dense packing of the molecules and their high dipole moments result in high oscillator strengths that make the bacteria (and organic matter in general) good candidates for strong coupling between excitons and photons. Their size, approximately 2 μm × 500 nm, means that they can also be inserted into a micron-sized optical microcavities with well-defined photon mode energies. The strong coupling condition is met when the leakage of the light trapped in the microcavity is slow compared to the energy exchange rate between the light and bacteria. This results in a modification of the energy spectrum of the exciton and cavity modes, with new energy levels corresponding to excitations called polaritons, arising from the coupling of the excitons and light. In [3] the transmission spectrum of the bacteria coupled to the cavity modes was probed with white light (see experimental setup in the figure 1).

Figure 1. Scheme of the experiment with strongly coupled living bacteria and light, [3]. The dye molecule (Trypan blue) is repelled by the bacteria during the duration of the experiment, providing a witness that they are alive.
Download figure:
Standard image High-resolution image{kind=link}
This resulted in two peaks, corresponding to the polariton branches, which have lower and higher energies compared to the uncoupled exciton energies. The so-called Rabi splitting is the energy difference between the two peaks when the uncoupled cavity mode and the exciton mode would have the same energy (zero detuning). The criterion for strong coupling is that the Rabi splitting is greater than the sum of the uncoupled photon and exciton energies; it is equivalent to the energy exchange rate being faster than the system dephasing. This condition was satisfied in the experiment, [3]. We shall now discuss a fully quantum-mechanical model for the formation of the polariton branches, which explains their existence and the Rabi splitting via entanglement between the bacteria and cavity EM field. The amazing fact is that the bacteria can actually stay alive during the experiments, despite the fact that they are strongly coupled to the cavity light. (The test of being alive in [3] was a form of homeostasis—the fact that the bacteria repelled a certain dye molecule during the experiment, which they absorb when dead.)
Quantised model
In our model, the BChl molecules situated in living bacteria are modeled as electric dipoles and reduced to two-level systems. Each of these dipoles is strongly coupled to the single frequency of light in the cavity through the Jaynes–Cummings Hamiltonian. The bacteria are modeled as a system of N two-level atoms, collectively interacting with the field of a single-mode cavity whose annihilation (creation) operator is a† (a). Each two-level atom is modeled as a pseudo-spin whose Pauli spin matrices are  . We assume a realistic small-sized bacterial sample and neglect the variations of the cavity field at its location. The BChls tend to lie along the axis of the aggregate. The number N of coupled dipoles in the sample assuming complete alignment can be estimated at about 95 million [6]. The coupling strength would in general depend on how many dipoles are within the region being probed by the light. In our simple model we assume an average coupling constant g between each of the dipoles and the field. This is a good assumption given that the wavelength of light is larger than the spatial extent of the sample. As we said, in [3] the strong coupling condition is satisfied, which means that the rate of energy exchange with the field is larger than the rate of dephasing. The individual g has never been directly measured for the bacterial sample, nor the individual dephasing rate. We therefore assume that for our model g is in the range that fits the observed collective Rabi splitting, so that it satisfies the strong coupling condition.
. We assume a realistic small-sized bacterial sample and neglect the variations of the cavity field at its location. The BChls tend to lie along the axis of the aggregate. The number N of coupled dipoles in the sample assuming complete alignment can be estimated at about 95 million [6]. The coupling strength would in general depend on how many dipoles are within the region being probed by the light. In our simple model we assume an average coupling constant g between each of the dipoles and the field. This is a good assumption given that the wavelength of light is larger than the spatial extent of the sample. As we said, in [3] the strong coupling condition is satisfied, which means that the rate of energy exchange with the field is larger than the rate of dephasing. The individual g has never been directly measured for the bacterial sample, nor the individual dephasing rate. We therefore assume that for our model g is in the range that fits the observed collective Rabi splitting, so that it satisfies the strong coupling condition.
Then, we introduce the total angular momentum J of the atomic sample with components  and
and  and consider time-independent single-spin energy splittings ωb. The Hamiltonian of the system in the dipole approximation thus reads (with ℏ = 1 throughout)
and consider time-independent single-spin energy splittings ωb. The Hamiltonian of the system in the dipole approximation thus reads (with ℏ = 1 throughout)

where ωa is the frequency of the cavity and g is the vacuum Rabi frequency (ignoring the vacuum contribution without any loss of generality and assuming ℏ = 1). The parameter j is the cooperation number in the Dicke theory [7], that is an eigenvalue of J2 which, together with the eigenstates of Jz, is used to build the Dicke states. The ensemble of N two-level atoms is then described as a pseudo-spin of size j = N/2. We can now apply the Holstein-Primakoff transformation, defined as
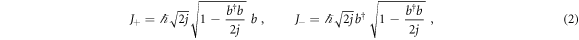
and

In the limit j ≫ 1 (thermodynamic limit) we obtain
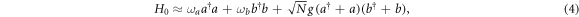
The eigenstates of this Hamiltonian are typically entangled states between a number of excitations in one of the oscillators (representing excitons in the bacteria) and the other harmonic oscillator (representing photons in a single mode of light), see [8]. Such 'dressed' states between excitons and photons are known as polaritons. These quasi-particles have a mixed physical character that is part-matter and part-light, and are responsible for the Rabi splitting observed in [3]. The dense packing and high oscillator strengths in organic media make them well suited to giving rise rise to these quasi-particles via strong coupling with light. A special class of these particles, known as organic surface plasmon polaritons, have been showcased in a series of recent experiments where quantized light and organic matter were strongly coupled via cavities and organic semiconductors [9, 10], down to the level of manipulating single organic molecules at room temperature [11]. Besides the inherent interest that the hybrid states of light and matter present, they are predicted to find uses such as in controlling the chemical kinetics in a wide class of photochemical reactions [12].
To demonstrate entanglement we use entanglement witnesses, which are physical observables that 'react' differently to entangled and disentangled states. Here we will only focus on ground-state entanglement since the relevant frequencies are at least one order of magnitude larger than kT at room temperature at which the experiment was carried out. In our case of two effective coupled harmonic oscillators we can use the energy as a witness. For simplicity, we assume that ωa = ωb = ω, i.e. that the exciton frequency is on-resonance with the light in the cavity. The energy of separable states of the form  can never be smaller than
can never be smaller than

The ground state of H0 the energy is lower by the vacuum Rabi splitting  (we are ignoring the vacuum contribution without loss of generality), [8]. Hence the ground state is an entangled state. We can therefore conclude from the measurements of Rabi splitting on two coupled harmonic oscillators assumed for light and bacteria (treated as a large collection of dipoles), that they are entangled. Specifically, the entangled subsystems are the excitons in the bacteria and the photons in the cavity. This concludes our analysis of the Rabi splitting observed in [3] using a fully quantised model for both dipoles and EM field. We now turn to discussing a few important points.
(we are ignoring the vacuum contribution without loss of generality), [8]. Hence the ground state is an entangled state. We can therefore conclude from the measurements of Rabi splitting on two coupled harmonic oscillators assumed for light and bacteria (treated as a large collection of dipoles), that they are entangled. Specifically, the entangled subsystems are the excitons in the bacteria and the photons in the cavity. This concludes our analysis of the Rabi splitting observed in [3] using a fully quantised model for both dipoles and EM field. We now turn to discussing a few important points.
Discussion
The entanglement witness above is also effectively a measure of entanglement. Namely, the ground state is entangled so long as g > 0. The amount of entanglement between bacteria and light could be quantified by the linear entropy of either of the subsystems. The linear entropy is defined as the complement of the purity of the state, i.e., 1 − Tr{ρ2} where ρ is the reduced density operator of either of the subsystem (obtained by tracing out the degrees of freedom of the other subsystem). A simple calculation shows that the linear entropy is  , where
, where  . In [3] the highest coupling achieved was
. In [3] the highest coupling achieved was  and therefore the amount of entanglement measured was 0.04.
and therefore the amount of entanglement measured was 0.04.
A question could be raised as to whether there might be other processes that could affect this entanglement. For instance, the cavity could leak photons which could spoil the coherence of polaritons. However, we take it as an observational fact that polaritons last long enough to be detected and must therefore preserve coherence during this period. The Q factor of the cavity used in [3] is 25, which corresponds to a photon lifetime of about 10 fs. Hence, this sets the relevant time-scale against which comparing all the other possible sources of decoherence.
Excitons could also dissipate within bacteria through interactions with phonons, for instance, but these processes are typically on much longer timescales than the cavity photon lifetime. In particular, the chlorosome exciton lifetime is on the order of 1 ns—much longer than the cavity photon life-time. In addition, the exciton on the chlorosome could migrate to the next energy level inside the living bacterium, but that transition is much faster (about 12 ps [13]). Therefore we believe that our model, albeit simple, is sufficient to describe the experiment.
It is worth pointing out that the same results (namely the observation of the vacuum Rabi splitting) can equally well be explained by a completely classical analysis, see for example [14–16]. There is no contradiction with the entanglement witness in our model, because the witness already assumes that both systems are quantum and checks whether the subsystems are entangled. In the classical analysis, on the other hand, the cavity mode is represented by a simple harmonic oscillator and is coupled to N classical dipoles each of which represents a BChl molecule within the bacteria. The emission spectrum obtained via the multi-beam interference technique (assuming that the cavity mode is on resonance with the dipole frequency) [14] exhibits a splitting in the intensity peaks separated by the classical analogue of the vacuum Rabi splitting, given by:
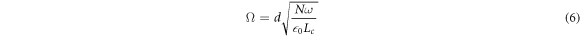
where  0 is the vacuum permittivity, Lc is the length of the cavity, and d is the dipole moment. This classically obtained result is exactly the same as that obtained by the full quantum analysis above (
0 is the vacuum permittivity, Lc is the length of the cavity, and d is the dipole moment. This classically obtained result is exactly the same as that obtained by the full quantum analysis above ( where g is the vacuum Rabi frequency) and, interestingly, this also includes the square root scaling with the number of oscillators (i.e. the molecules).
where g is the vacuum Rabi frequency) and, interestingly, this also includes the square root scaling with the number of oscillators (i.e. the molecules).
One important subtlety should be mentioned here—relating to the indirect inference of entanglement in our system. Suppose that instead of the fully classical model, or indeed the fully quantum model, we use a semi-classical model. By semi-classical model we mean that the bacteria are treaded as classical harmonic oscillators, while light inside the cavity is treated as a quantum system—i.e. as a field of quantum harmonic oscillator. Note that more conventionally a semi-classical model would treat the bacteria as quantum, and the light as a classical field; but for present purposes, the former case is of interest because our model is about showing that the bacteria are quantum dipoles. In any case, neither of the two semiclassical models can obtain the vacuum Rabi splitting. This is a special case of a more general fact that semi-classical models, as defined above, can never fully reproduce quantum features—see for example, [17]. The fact that the semi-classical model fails to reproduce certain correlations between two subsystems is not unusual; in fact, it also features in cryptographic protocols, [18], where four bits can reproduce some correlations of a singlet, but one qubit and two bits cannot.
The fact that a semiclassical model is ruled out by the Rabi splitting hints to an interesting speculation. Light is known to be a quantum field from other independent experimental evidence, so a non-approximative model for light would require it to be quantum. As soon as light is treated as quantum, since the Rabi splitting is observed in the experiment in [3], the bacteria (more precisely, whatever degree of freedom within bacteria couples to light) would also have to be quantum—in the sense that they must be described by a model with at least two non-commuting observables. This is because, unlike for other simpler light-harnessing phenomena such as photon absorption, the Rabi splitting cannot be explained by a semi-classical model where the field is quantum and the other system is classical.
The witness of non-classicality provided by the Rabi splitting is however not enough to rule out hidden-variable models. There are indeed more refined and direct levels of probing and confirming the degree of entanglement between two subsystems, here being the two harmonic oscillators, one representing the bacteria and the other the cavity mode. At the most detailed level we have Bell's inequalities [19], which would indeed rule out a hidden-variable model for the composite system of bacteria and light. To detect a violation of the inequality, one would need to probe the bacteria and light separately, by measuring two suitably chosen conjugate observables for each. For instance, one could measure (effective) x and p on both the dipoles in the bacteria and the cavity light. This is of course much harder to achieve experimentally. Harder still would be to spatially separate the bacteria and light to ensure that there is no signal propagation between the two while measurements are carried out (to ensure that the locality loophole is closed). Another possible line of probing the quantum character of the bacteria-light interaction is to investigate the quantum coherent dynamics of polaritons. A less challenging experiment would be the observation of Rabi flopping between the two Rabi split levels associated with the polariton. In this case, the light used to excite the cavity would also have to be coherent, unlike the experiment analysed here. However, given that the spectrum obtained in [3] is effectively just the modulus squared of a Fourier Transform of the Rabi flopping, it is still questionable whether this would be a conclusive proof of quantum coherence, while it would certainly not be a witness of entanglement since the bacteria and light are treated as one quantum system, namely a polariton. At the harder end of the spectrum we also have a violation of the Leggett-Garg and related inequalities (e.g. [20]). This would again require identifying suitable observables to be measured on the polariton, as well as devising a means of performing a non-invasive (in practice, weak) measurement.
Acknowledgments
The Authors thank three anonymous referees for helpful suggestions and comments. VV and TF thank the Oxford Martin School, Wolfson College and the University of Oxford, the Leverhulme Trust (UK), the John Templeton Foundation, the EU Collaborative Project TherMiQ (Grant Agreement 618074), the COST Action MP1209, the EPSRC (UK) and the Ministry of Manpower (Singapore). This research is also supported by the National Research Foundation, Prime Ministers Office, Singapore, under its Competitive Research Programme (CRP Award No. NRF- CRP14-2014-02) and administered by Centre for Quantum Technologies, National University of Singapore.