
Abstract
Adult tissue stem cells can form self-organizing 3D organoids in vitro. Organoids resemble small units of their organ of origin and have great potential for tissue engineering, as well as models of disease. However, current culture technology limits the size, architecture and complexity of organoids. Here, we review the establishment of intestinal and hepatic organoids and discuss how the convergence of organoids and biofabrication technologies can help overcome current limitations, and thereby further advance the translational application of organoids in tissue engineering and regenerative medicine.
Export citation and abstract BibTeX RIS
Introduction
The establishment of three-dimensional (3D) organoid cultures from multiple organs has been a major breakthrough and has gained increasing attention over the last ten years. A historical perspective of the evolvement of 3D tissue cultures, and the different definitions of the term organoids has recently been reviewed [1]. In general, organoids are self-organizing 3D structures that grow in vitro, embedded in an extracellular matrix (ECM) and resemble their organ of origin [2]. They can be established from different cell sources, such as primary tissue explants, cell lines, multipotent adult stem cells, pluripotent embryonic stem cells (ES cells) or induced pluripotent stem cells (iPS cells), and from a variety of tissues [1, 3]. In this perspective, we focus on intestinal and hepatic organoids from adult stem cells or iPS cells, and how the integration with advanced biofabrication technologies may enhance the utility of organoids in research and therapeutic applications.
The generation of 3D organoids stems from our increasing understanding of stem cell signaling pathways [4]. Key components of stable hepatic and intestinal organoid cultures include agonists of WNT and epidermal growth factor, antagonists of bone morphogenetic protein (BMP), and embedding the cells in an ECM. Under those culture conditions, adult stem cells [5, 6] can produce all progeny of their tissue lineage and self-organize into small 3D structures closely mimicking the homeostasis and function of their native organ [7]. Organoids can also be derived from pluripotent iPS or ES cells, which can be specified into definitive endoderm, and subsequently into a hindgut [8, 9] or hepatic fate [10, 11]. Hindgut spheroids form intestinal organoids when cultured in the same conditions as adult intestinal stem cells, containing all intestinal epithelial cell types with mesenchymal cells surrounding the organoids [8, 9]. When cultured in expansion medium with active WNT signaling, intestinal and hepatic organoids appear as hollow cyst structures and are highly proliferative. In differentiation conditions, where WNT signaling is blocked, organoids become smaller and cell-dense and upregulate the expression of differentiation markers [4–6, 12] (figure 1). Since the stem cells retain their self-renewing capacity in culture, organoids represent an unlimited source of primary epithelium, which has been shown to remain genetically stable [13], especially for organoids derived from adult stem cells [5, 6, 14]. Hence, organoids are already widely applied in in vitro studies [2, 7], for example as disease models, and hold great potential for transplantation and tissue regeneration purposes [15–18].

Figure 1. Schematic representation of human organoid cultures from adult tissue stem cells. Progenitor or stem cells are isolated from a hepatic or intestinal biopsy, respectively. Single isolated cells are cultured in Matrigel and expansion medium, where they form cyst-like structures. Hepatic organoids consist of stem cells (dark blue) and ductal progenitor cells (light blue). Intestinal organoids mainly consist of stem cells (green), Paneth cells (yellow) and transit amplifying (TA) progenitor cells (brown). When exposed to differentiation medium, hepatic organoids become denser and contain ductal cells (light blue) and hepatocyte-like cells (red). Differentiated intestinal organoids display a folded structure and contain all differentiated epithelial cells of the small intestine (enterocytes, goblet cells, enteroendocrine cells and tuft cells, depicted in light pink, purple, dark red and orange, respectively).
Download figure:
Standard image High-resolution imageDespite their great potential, current organoid culture conditions feature several limitations that prevent accurate recapitulation of tissue architecture and in vivo behavior. Matrigel, which is often used, is an undefined ECM of murine origin, which hampers translation to humans due to immunogenicity. Local signaling gradients, in vivo, tightly control cell fate and tissue architecture; in contrast, organoid cultures offer no spatiotemporal control over the provided growth factors. Moreover, organoids are restricted in size due to limitations of nutrient diffusion, and lack complex interactions with non-parenchymal cell types. Consequently, the architectural organization, maturation status and functionality of organoids does not yet reach in vivo levels despite the fact that they contain all differentiated epithelial cell types found in vivo [6, 19]. We anticipate that combining organoids and biofabrication technologies may overcome those limitations, and produce larger, functional tissues under highly controllable culture conditions.
Biofabrication aims to recapitulate tissues and organs by combining cells and biomaterials into very specific structures with dictated organization. Recently, biofabrication was defined as
...the automated generation of biologically functional products with structural organization from living cells, bioactive molecules, biomaterials, cell aggregates such as micro-tissues, or hybrid cell-material constructs, through bioprinting or bioassembly and subsequent tissue maturation processes [20].
Biofabrication effectively employs and combines an array of technologies, methods and material compositions. It includes direct cell-laden printing methods using inkjet printing [21] or robotic dispensing [22] as well as indirect or multi-stage methods, such as melt electrospinning writing [23], which allow us to simply shape, direct and promote the development of cell-material constructs.
Mimicking the ECM
The ECM regulates cell behavior and is an essential component of the stem cell niche. In vivo, hepatocytes are in close proximity to the ECM in the space of Disse, and intestinal epithelial cells face an underlying basement membrane. In the liver, receptors of the β1 integrin family interact with collagen type I, fibronectin and laminin that are present in the ECM [24, 25]. In the intestine, the basement membrane is rich in collagen type IV and laminins, which provide adhesion sites for receptors of the integrin superfamily that are expressed on the basolateral membrane of intestinal epithelial cells [26–28]. The ECM not only anchors the epithelial cells, but also influences proliferation, differentiation and migration of the interacting cells [27, 29, 30]. Differentiation of liver progenitor cells towards the hepatocyte or cholangiocyte lineage [31] and proliferation and differentiation of intestinal stem cells [32] are not only regulated through biochemical signals (the matrix composition), but also through the mechanical properties (stiffness) of the ECM. Mimicking the biochemistry and biomechanics of the native ECM of a certain tissue in vitro is thus crucial for controlling proliferation and differentiation of the cells that constitute an organoid.
Hepatic and intestinal organoids are currently mostly cultured in Matrigel [4–6, 12, 33], a gelatinous protein mixture with high biological activity that is secreted by mouse sarcoma cells [34]. The protein complexity of Matrigel is advantageous as it closely mimics the composition of the basement membrane, and enables proliferation, differentiation and self-organization of embedded cells into 3D structures [35]. However, the murine origin, complex extraction process and undefined composition of Matrigel all lead to a relatively high batch-to-batch variation in composition and stiffness, and hamper in vivo applications in humans due to immunogenicity. Cellmatrix type I-A has also been used to culture intestinal organoids in one study [36]. The collagen-based gel is more defined than Matrigel, however the porcine origin still hampers clinical translation. Thus, in order to advance the maturation and translational potential of organoids to the clinics, there is a need for a defined hydrogel that can substitute and overcome the disadvantages posed by animal-derived gels.
The use of synthetic or biological hydrogels may bypass the need for Matrigel. Synthetic polymers are chemically defined and allow for a tight control over the mechanical properties, however, they lack biofunctionality, and thus require the incorporation of biomolecules to recapitulate the biological functions of the ECM [37]. Biological polymers, such as gelatin, are biofunctional, but the mechanical properties are more difficult to control. In recent years, biomaterial approaches have focused on the physicochemical tailoring of biological hydrogels to control their mechanical properties. This is of particular importance when applied to the culture of organoids since these are sensitively regulated through mechanotransduction [38]. The mechanical properties of hydrogels can be fine-tuned by varying the mixing ratio of their components [39], or by the reinforcement of very soft gels with more mechanically and dimensionally stable structures [40].
Gelatin-methacryloyl (GelMA) hydrogels, for example, are fabricated by the addition of methylacryloyl side groups to gelatin. This process makes gelatin crosslinkable, and thus yields a hydrogel that is biofunctional and mechanically tunable [41, 42]. It is highly versatile and suitable for processing, since it can be easily crosslinked by either enzymes or light, which makes it suitable for direct 3D printing [41]. Likewise, we recently showed that enzymatically crosslinkable poly-ethylene glycol (PEG) hydrogels can be functionalized with a covalently linked Arg–Gly–Asp peptide (which represents binding to fibronectin) to support the expansion of intestinal stem cells [32]. Subsequently, by varying the mechanical properties and decreasing the stiffness of the functionalized PEG hydrogel in a modular manner, differentiation and polarization of intestinal epithelial organoids could be induced [32]. Thus, by mimicking the biological and mechanical ECM properties of a particular tissue with a tailor-made and defined hydrogel, proliferation and maturation of organoids can be controlled and improved. These defined and modular hydrogels offer new possibilities to study the influence of biological and mechanic properties of ECM on cell behavior in the self-organizing organoid system. In addition, synthetic hydrogels can be altered for a particular application. For example, organoids can be cultured in a functionalized hydrogel with a relatively high stiffness that promotes proliferation, when large amounts of organoids are needed. In contrast, the use of modular hydrogels for the culture of organoids may increase their maturation status and functionality. Additionally, the defined nature of synthetic hydrogels may provide exciting opportunities for the translation of organoid technology to clinical settings.
Complex scaffolds to mimic tissue architecture
The small intestine is highly structured. The epithelium is organized into proliferative crypt compartments which are embedded in the intestinal submucosa, and extruding villi harbor the differentiated cells of the small intestinal epithelium [43]. The proximal duodenum contains a high number of very long villi, whereas the abundance and lengths of the villi decreases towards the distal part of the small intestine [44]. This defined topography affects the mechanics and fluid dynamics of the small intestine in vivo [45]. The importance of topography on cell fate decisions can be demonstrated by intestinal Caco-2 cells, which differentiate along the crypt-villus axis in vitro when seeded on a collagen scaffold that reflects the shape and size of villi [46].
In current organoid culture conditions, both murine and human small intestinal organoids do not contain villus architecture. Murine small intestinal organoids self-organize into crypts and flat villus-like domains, however do not build real extruding villi [4]. Human small intestinal organoids are cultured as round cysts under expansion conditions, and undergo a change in morphology into a highly folded structure upon exposure to differentiation medium [5, 14], with an even more pronounced lack of clear villus architecture.
Biofabrication techniques, such as laser ablation or stereolithography, using bioinert photopolymer resins, allow easy and fast production of a scaffold with villus topography [47]. Such a scaffold can be either directly seeded with cells, or used as a mold for the subsequent production of a hydrogel-based (porous) scaffold [48]. Indeed, murine and human intestinal organoids were recently grown on such a scaffold in vitro and after omental implantation into immune-deficient mice in vivo, where they supported the growth of epithelial cells and blood vessels [49]. As such, the combination of hydrogel-based scaffolds and organoids may be the next step in the development of a tissue-engineered small intestine for the treatment of patients with short-bowel syndrome and other intestinal diseases.
Flow perfusion culture for homogeneous distribution of nutrients and oxygen
In order to create larger tissues composed of multiple organoids, flow perfusion bioreactors may bypass the limitations of size, oxygen supply and nutrient/waste transport. These bioreactors enable perfusion of a biofabricated tissue or tissue construct; support shear stress by microfluidics, which can maintain or even increase function of parenchymal cells [50]; provide a uniform medium distribution, and maintain the desired concentration of gases and nutrients in the medium [51]. Flow perfusion bioreactors have shown great potential over previous bioreactor systems, such as spinner flasks and rotating wall vessels [52]. All flow perfusion bioreactors consist of a similar setup with a pump, tubing, media container and a perfusion chamber. The design of the perfusion chamber is customized to the tissue of interest and intended application.
These flow perfusion bioreactors provide a highly controlled system for bioengineered tissues. For instance low shear stress can polarize hepatocytes in a bioreactor, thereby increasing the function of drug-metabolizing cytochromes [53–56]. For intestinal tissue models using organoids, flow perfusion bioreactors have been developed which allow long-term perfusion and polarized transport from the apical side of the epithelial cells towards their basolateral side [57] and increase functions, such as multidrug resistance protein 1 (MDR1) transporter activity [58]. In addition, a more homogeneous maturation status of cells within organoids may be achieved if growth factors could reach the cells more easily by using porous tissue constructs. Porous tissue constructs in flow perfusion bioreactors would also increase the distribution of oxygen and nutrient/waste transport and provide a better and more constant exposure. Taken together, flow perfusion bioreactors will prove vital in maintaining tissue-specific functionality and providing proper distribution of gases and nutrients in the media.
Spatio-temporal control over growth factors
Current organoid culture systems do not regulate the spatial and temporal signaling cascades (WNT, Notch, transforming growth factor beta, BMP and hedgehog) necessary for homeostasis and regeneration in the intestine [43, 59] and liver [60]. A range of growth factors are added to the culture medium to mimic the in vivo signaling pathways, however all cells are exposed to a uniform concentration of growth factors, whereas concentration gradients in vivo are crucial for localized cell fate decisions. As such, human intestinal and liver organoids mainly consist of stem and progenitor cells under expansion conditions, and require the withdrawal of several growth factors in order to induce differentiation [5, 6].
Biofabrication will allow us to expose organoid cells to their required growth factors in a modular, highly controllable and reproducible manner [61]. Spatio-temporal control over growth factors and small molecules can be achieved by using specific biomaterials, which are able to either define the length of the release period [62] or promote release of bioactive molecules in multiple discrete stages [63, 64]. Other more complex strategies can use materials that release drugs, on demand, as a response to specific external stimuli such as local pH [65], temperature or combinations thereof [66].
Patterning of organoids and non-parenchymal cells
In both the intestine and liver, non-parenchymal cells heavily influence tissue homeostasis and regeneration. Intestinal [4, 5, 14] and hepatic [6, 12, 33] epithelial organoids from adult tissue stem cells are a simplified model of the complex in vivo environment and lack non-parenchymal cells that intersperse and interact with the epithelial cells in vivo. The simplistic nature of organoids is an advantage when using a reductionist approach to studying stem cells and their progenies, without the confounding influences of their (cellular) environment. However, the lack of a native microenvironment hampers studies involving immune responses and interaction of epithelial cells with their supporting cells, and might limit the maturation of epithelial cells due to missing instructive cues.
In the intestine, the epithelium is in close spatial proximity to several non-parenchymal cells, including the underlying mesenchyme and immune cells. Mesenchymal cells have been shown to drive villus formation [67], and secrete growth factors to the epithelium. For example, intestinal epithelial stem cells receive niche signals from neighboring Paneth cells [68], but upon ablation of Paneth cells in a mouse model, the stem cells remain functional [69] due to redundant WNT ligands secreted by surrounding mesenchymal cells [70]. In vitro culture of Paneth cell-deficient Atoh1-/- KO organoids on an intestinal mesenchymal layer rescued their survival [70], and culture of small intestinal wildtype organoids on a layer of stromal cells [71, 72] or subepithelial myofibroblasts [73, 74] promoted organoid formation and survival without addition of exogenous WNT agonists. In addition, innate immune cells have been shown to support stem cell survival and organoid growth by the secretion of IL-22 [75].
The liver consists of the parenchymal hepatocytes and several non-parenchymal cell types such as liver mesenchymal stromal cells, sinusoidal endothelial cells, stellate cells and Kupffer cells. When cultured on a fibroblast feeder layer, adult hepatocytes show an increased in vitro proliferation capacity [76]. Kupffer cells, the tissue resident macrophages of the liver, play a pivotal role in tissue homeostasis, and seem to facilitate liver injury by the secretion of inflammatory cytokines and reactive oxygen species [77]. Furthermore, the sinusoidal endothelium promotes liver regeneration by the secretion of hepatocyte growth factor and angiocrine signals [78, 79].
We expect that biofabrication will help create the complex cellular microenvironments found in vivo by enabling the patterning of several cell types with high spatial accuracy. Specialized software allows precise and easy design of heterogeneous constructs, which can define various patterns, gradients and densities of cells [80] and material compositions [81]. In addition, the required biochemical and mechanical hydrogel properties may vary between cell types within one tissue, and greatly influence the behavior of developing tissues. To account for those different local requirements, multiple types of cell-laden gels can be combined through robotic dispensing technologies [82] or by melt electrospinning writing [83] for gels with more solid structures. A good example of the importance of cellular patterning can be found in liver applications, where bioprinting of hepatocyte progenitor cells (HPCs) and non-parenchymal cells in a 3D model with hexagonal architecture was found to improve morphological organization and liver-specific functions of the HPCs [84]. This kind of patterned combination of cell-laden materials can result in the generation of constructs composed of multiple inner biomechanical microenvironments, which may be specifically suitable for organoids and different non-parenchymal cells.
Vascularization of organoids and tissues
One of the major challenges for the biofabrication of larger multi-organoid structures (or even organs) is vascularization [85]. Diffusion for delivery of nutrients, oxygen, growth factors, and removal of metabolic waste is often already insufficient when tissue thickness exceeds 100–200 μm [86], and organoids easily reach sizes above 300 μm. Diffusion in a bioengineered tissue can be achieved with well-distributed and interconnected vascular networks. Several approaches that can create neovascularization appropriate for diffusion include the incorporation of endothelial cells with scaffolds; in layered cell sheets by 3D printing; in co-culture with spheroids or aggregates of parenchymal cells; or with the addition of angiogenesis-inducing growth factors [87, 88]. One promising approach using 3D bioprinting creates channels with a sacrificial material, e.g. agarose or Pluronic F127, enclosed inside a cell-laden hydrogel matrix. After removal of the sacrificial material, these microchannels can be filled with endothelial cells [89–91]. Patent lumens are thus created with endothelial linings, allowing better representation of native tissue such as liver [92].
Endothelial cells are essential for vascularization and angiogenesis, and they communicate with smooth muscle cells to promote and stabilize vascular development and function [93]. Human umbilical vein endothelial cells are often used in these types of co-cultures as a source for endothelial cells. However, the type of endothelial cells required depends on the application and tissue of interest [85]. For example, capillaries in the intestine have a continuous endothelial lining whereas capillaries in the liver are leaky and have large intercellular gaps [94]. The addition of endothelial cells in bioengineered liver tissue using organoids allows vascularization and transplantation [16, 95], but ex vivo bioengineered liver tissue based on organoids might also benefit from the addition of endothelial cells by increasing hepatic function [96, 97]. Addition of cells that promote or stabilize vascularization can also be achieved by adding (vascular) smooth muscle cells [93]. As organoids easily reach sizes above 300 μm, vascularization is a necessity for the use of organoids for tissue engineering approaches. Although vascularization is still considered one of the greatest challenges in tissue engineering, methods such as the use of sacrificial material combined with endothelial cells in organoid-laden hydrogels will lead to bioengineered tissues that better represent in vivo physiology.
Conclusions
Organoids represent an exciting new organ model, since they can be established from a variety of species, resemble their organ of origin, and can be infinitely expanded in culture while retaining their genetic integrity. Current limitations of organoids are caused by the lack of fully controllable culture conditions, and therefore, the inability to reproduce the microenvironments necessary for the proper behavior and organization of cells; to induce vascularization; and to control cell–cell and cell–matrix interactions. Biofabrication offers a wide variety of techniques that can introduce these additional dimensions of control to organoid cultures (figure 2).

{kind=link}
Figure 2. Improvement of organoid technology by convergence with biofabrication. Depicted are 5 examples: (1) Bioprinting of a porous tissue structure by the incorporation of a sacrificial hydrogel allows perfusion culture of printed organoid constructs. (2) Biological and mechanical properties of hydrogels can be spatially adjusted for optimal proliferation and differentiation of organoid cells. (3) Spatial control over organoids and non-parenchymal cell types by bioprinting technologies. (4) The incorporation of endothelial cells can lead to vascularization and increased size of organoids. (5) A hydrogel scaffold mimicking in vivo topography can be seeded with organoid cells for improved architecture and differentiation. Further details are provided in the text.
Download figure:
Standard image High-resolution image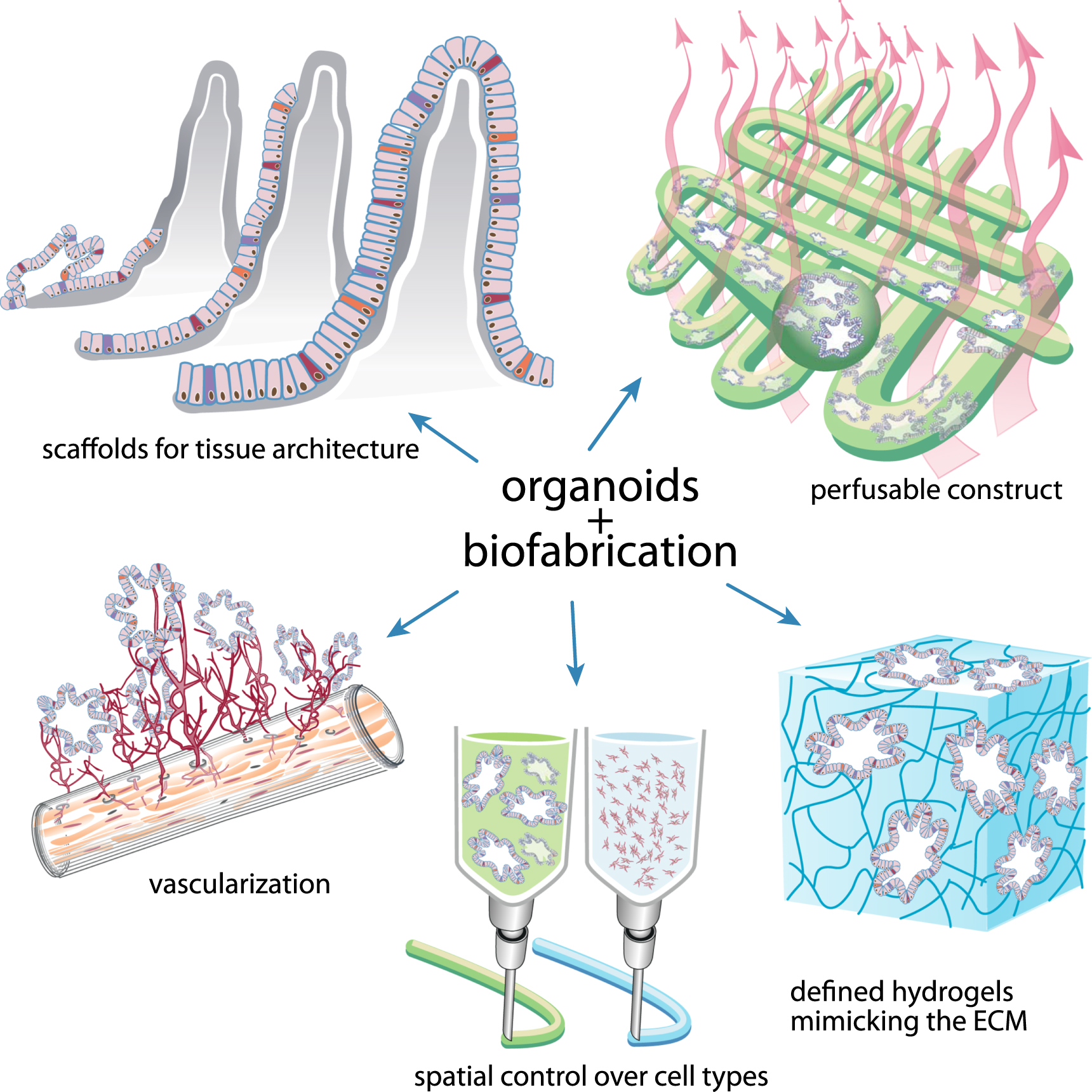{kind=link}
We envision an additive effect of biofabrication on the versatility of organoid technology. For example, the use of a defined synthetic hydrogel with control over biological cues will increase spatial control over proliferation and maturation of organoids in a tissue-specific manner, while also potentially enabling translation of organoids towards the clinic. Biofabrication of scaffolds on which organoids can be seeded will allow the growth of a tissue with in vivo-like architecture, which may increase maturation and functionality of organoid cells and could potentially be used for transplantations in the future. Advances in biomimetic hydrogels and 3D bioprinting will allow us to combine organoids and supporting cells with highly defined spatial control. The culture of organoids or organoid-derived porous tissue constructs in a flow-perfusion bioreactor will increase the control over growth factor concentrations, oxygen and the pH in the medium, thereby facilitating a homogenous organoid culture with increased functionality. Finally, biofabrication will enable the incorporation of structural microchannels into organoid cultures, which can be seeded with endothelial cells in order to receive vascularized multiple-organoid constructs that will allow the formation of larger tissues without a necrotic core. In summary, we predict that the combination of biofabrication technologies and organoids will increase the organization, complexity and maturation of organoids, and thereby enhance their application potential, especially for tissue engineering and regenerative medicine.
In order for the convergence of biofabrication and organoid technologies to become a reality, it is necessary to define, fine-tune and standardize this modified approach. It will be challenging to faithfully support organoid biology with technically feasible methods, as the mechanical properties of tissue-engineered constructs are often a compromise between printability and biological performance [40]. In addition, building a multi-layer, multi cell type, complex 3D structure that mimics in vivo architecture and interactions (both internal and external) leaves each individual system open to a range of variables, and optimal conditions for each case will need to be established.
Nevertheless, both organoids and biofabrication have already proven themselves as great tools for regenerative medicine, and the combination will offer unmatched possibilities for creating complex and functional tissues that more closely resemble the in vivo situation.
Acknowledgements
All authors declare no conflict of interest. JM was supported by the Dutch Arthritis Foundation (LLP-12), the European Community's Seventh Framework Programme (FP7/2007-2013) under grant agreement n°309962 (HydroZONES) and the European Research Council under grant agreement n°647426 (3D-JOINT). BS was supported by the Dutch Research Counsel NWO ZON/MW (92003538 and 16004121).