
Abstract
The results of many medical measurements are directly or indirectly influenced by the autonomic nervous system (ANS). For example pupil size or heart rate may demonstrate striking moment-to-moment variability. This review intends to elucidate the physiology behind this seemingly unpredictable system.
The review is split up into: 1. The peripheral ANS, parallel innervation by the sympathetic and parasympathetic branches, their transmitters and co-transmitters. It treats questions like the supposed sympatho/vagal balance, organization in plexuses and the 'little brains' that are active like in the enteric system or around the heart. Part 2 treats ANS-function in some (example-) organs in more detail: the eye, the heart, blood vessels, lungs, respiration and cardiorespiratory coupling. Part 3 poses the question of who is directing what? Is the ANS a strictly top-down directed system or is its organization bottom-up? Finally, it is concluded that the 'noisy numbers' in medical measurements, caused by ANS variability, are part and parcel of how the system works. This topical review is a one-man's undertaking and may possibly give a biased view. The author has explicitly indicated in the text where his views are not (yet) supported by facts, hoping to provoke discussion and instigate new research.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Introduction
In the medical and research world for a long time there has been a strange aura around the autonomic nervous system (ANS): it was not 'dignified' like the neural motor control or the optic system. It is poorly identifiable, has a strange central and peripheral anatomy and is about bodily functions that, generally, are dismissed as 'vegetative'. This includes control of the gut and urine voiding, getting an erection or not; important, yes, but not the functions one likes to talk about.
The patients in the waiting room of the GP might tell one differently: a large proportion of the complaints that brought them there is due to or at least strongly related to some autonomic dysfunction. Still, the neurologist who specializes in autonomic functions and their derailment was considered a looser by his peers in the Neurology department. This all might just be changing with the advent of new insight from the lab and new diagnostic options in the clinic.
This Introduction into ANS function is intended for the (bio-)physics community to give a brief overview of autonomic functions and the biological hardware behind those. It is not intended to supplant standard physiology or anatomy textbooks; some level of basic knowledge will briefly be recapitulated. Next it will be shown that reality does not fit the straight forward textbook schematics. In particular, the extent of branching connections, the diversity of so-called co-transmitters and the high degree of organization in networks within the tissues rather than in more central neuronal structures, all call for a re-evaluation of how the biological machinery might work as a 'system'. Since this Introduction has only one author, it presents undoubtedly a biased view. Therefore I have adopted a system from Burton (1972) to mark those passages that may be considered controversial, or are not supported by sufficient experimental proof (yet) be marked by brackets as follows: [►.......◄]. A comprehensive review of basic knowledge of the ANS can be found in Dr Jänig's life work (Jänig 2006) and in Squire et al (2008) and Purves et al (2004); of the recent books which cover more the clinical and pharmacological aspects, (Low 2008, Mathias and Bannister 2013, Robertson et al 2012) should be mentioned.
Below is an overview of how the subject will be treated, divided into 3 chapters.
- 1.The peripheral autonomic nervous system, review of the basics.
- 1.1.Sympathetic versus parasympathetic system: anatomical and functional distinctions
- 1.2.Evolution and the 'smallest functional unit' of local organ control
- 1.3.Parallel innervation by the sympathetic and parasympathetic system
- 1.4.Function of ganglions and ganglionic plexuses; the enteric nervous system, how many 'little brains' do we have?
- 1.5.Transmitters and co-transmitters, pre- and post-ganglionic
- 2.Innervation of some specific organ systems, their reaction to sympathetic- versus parasympathetic activity.
- 3.The autonomic nervous system, orchestra or director?
- 3.1.Global versus local activity, what do measurements tell?
- 3.2.Central versus peripheral autonomic nervous system; a hypothesis.
- 3.3.Integration of autonomic and somatic nervous system; the immune system and the ANS.
- 4.Closing remarks
1. The peripheral autonomic nervous system, review of the basics
1.1. Sympathetic versus parasympathetic system: anatomical and functional distinctions
Observation and interpretation go often hand in hand. The first anatomical observations of what looked like a nerve with ganglia in the neck (in close proximity to the vagus nerve) and all along the spinal cord, sprouting its fine, unmyelinated branches to all internal organs, led to the idea of imposed 'sympathy' between even distant parts, hence the name 'sympathetic nerves'. The vagus nerve takes its name from the fact that like a wanderer its myelinated fibers can be traced from the brainstem, exiting the skull, to almost any place in the body. Where it does not reach, i.e. the lower abdominal and pelvic region, its function is taken over by myelinated nerves from sacral segments of the spinal cord. Functionally and anatomically, the two, sympathetic and vagus nerves were shown to have one point in common, which has become the hallmark of the autonomic system, i.e. the fact that no target organ is served directly from the central nervous system (CNS), but always a relay nerve cell outside the skull or vertebral canal is interposed. A conglomerate of such nerve cells is called a ganglion (plural ganglia). The ganglia of the sympathetic system were quite obvious: as shown in figure 1 for a section of the sympathetic trunk and its connections to the cord at the level just below the diaphragm. From the ganglia, thin, greyish, unmyelinated fibers emanate. The fact that the vagus nerve and other nerves that serve comparable functions also have ganglion cells interposed between nerve endings and target cells, went unnoticed for a long time. These nerves are a shiny white, well-myelinated, and the final ganglion cells are only found very close to the target organs. That is where the system draws its name from: the prefix 'para-' meaning: 'close to' or 'next to', hence: parasympathetic.
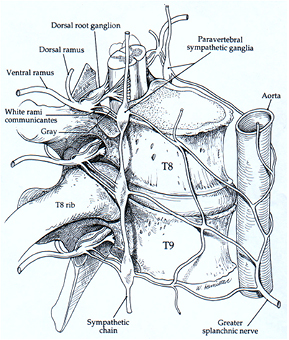
Figure 1. The paravertebral sympathetic chain ganglia at the level of the 8/9th thoracic vertebrae. Clearly shown are the 2 communicating branches with the spinal nerves which are protruding through the intervertebral foramina. Uninterrupted preganglionic fibers cross the ganglion and continue into a prevertebral plexus, where ganglia are formed around major branches of the aorta (below this level). Picture taken from Low, Clinical autonomic disorders, 1993, first edition, 2nd chapter by Y Harati on: Anatomy of the spinal and peripheral autonomic nervous system. Copyright Winifred J Hamilton, PhD, reproduced by permission.
Download figure:
Standard image High-resolution imageIt has taken a long time before the separation of what became known as the 'vegetative system' into two functionally separate systems became more or less clear. The one, broadly speaking associated with 'rest and recovery', is the parasympathetic system, importantly served by the vagus nerves, the 10th cranial nerves, with motor cells in the medulla oblongata. The other is the Flight, Fight, Fright system, served by the sympathetic trunk in a rope ladder system of ganglia next to and in front of the spinal column (terminology: paravertebral and prevertebral respectively), with motor cells in the spinal cord from the level of the lower cervical to the upper lumbar segments. If some 'central command' from higher brain centers requires (co)activation of the autonomic system (it generally does), sympathetic involvement must go via the spinal cord before a message is relayed to the ganglia where the transformation to commands to the peripheral organs takes place. This leads to unexpected consequences, like the signal for pupillary dilation, which originates from the level of the hypothalamus, but has to pass to the spinal cord and back via the sympathetic trunks into the head to find the muscles that dilate the pupils (pupillary reflexes are 'consensual' i.e. they occur in both eyes equally). One special part of the sympathetic trunk is the marrow of the adrenals (adrenal medulla): it consists mainly of so-called chromaffin cells, taking after their long-ago noticed property to oxidize the adrenaline they contain to a dark brown color under the influence of chromium. These cells are modified ganglion cells that do not have their own axons projecting to end organs, but they secrete their transmitter directly into the venous system, thereby spreading the sympathetic message to the whole body.
Finally, not implied in the enumeration, is the 'enteric nervous system' (ENS), which is an autonomic nervous system on its own, in function modulated but not regulated by the central parts of the ANS. This will be detailed in the paragraphs dealing with specific organs.
1.2. Evolution and the 'smallest functional unit' of local organ control
1.2.1. A digression into evolution.
The first section gave a 'top-down view' of the ANS as a specialized part of the central nervous system, extending its branches to the targets in the periphery. However, the further away from the center one goes, the stranger this 'system' becomes, as will be shown below. A better understanding may be obtained when the evolution is traced, as far as this is possible for functional systems: they have not been fossilized and their history may only be guessed from extant animal species. Alternatively, the embryological development may be followed if that is taken as a partly replay of evolution where quickly developing local demand forces the on- and off-switching of genes that determine intermediary and ultimate shape and function.
One of the 'simplest' organism that might be considered part of the line of forerunners of what eventually evolved into vertebrates is Hydra or the freshwater polyp. Broadly speaking it consists of an elongated gastrula ('little stomach'): comparable to the stage where the fertilized egg, after a series of cell divisions, gets an indentation that deepens and grows to what later will become the inside of the complete gastrointestinal tract, from mouth (the original opening of the indentation) to anus (which is a later invention in evolution). Like the gastrula, Hydra's body is composed of only two epithelial cell layers: an outer ectodermal and an inner endodermal layer. Around the mouth opening at Hydra's head region is a circular contraption, where tentacles are attached that catch prey by shooting off poison-filled harpoon-like nettles and help bring the food into the stomach body of the animal (see figure 2).
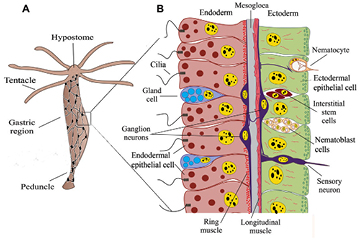
Figure 2. (A) Hydra vulgaris, showing its nerve net (total height about 10 mm). (B) Detail of the side wall, showing the inner (endodermal) and outer (ectodermal) epithelial cells. These are separated by the mesogloea, a cell free layer. Attached to the ectodermal cells are longitudinal muscle fibers. Figure slightly adapted from Technau and Steele (2011); reproduced by permission. (Nerve-net in (A) inspired by Hejnol and Rentzsch (2015).
Download figure:
Standard image High-resolution imageImportant for the evolution of motion and a nervous system are some specialized cell types present in Hydra's wall: myo-epithelial cells and primordial nerve cells. The myo-epithelial cells are part of the lining cell layers, but they have a contractile extension at their basal side extending into the gel-like cell-free layer between ecto- and endoderm. At Hydra's level of development, the distinction between autonomic and somatic nervous system does not exist. The nerve cells are concentrated in a ring around the mouth/head, then organized in a network around the body and there is a second concentration around the foot-part (peduncle) (Hejnol and Rentzsch 2015). This nervous system coordinates movements of the animal, for instance when entering a prey into the stomach or moving to a new place, functions which are difficult to get from just interconnected (myo-)epithelial cells. The nerve fibers run along their target cells in close apposition, but these have no developed receptive field, therefore there is no motor end plate. They have both clear and dense-core vesicles, filled with neurotransmitters. The first transmitters that have been identified were neuropeptides (Koizumi et al 1989), which are the oligopeptide molecules responsible for the dense coloring of the transmitter vesicles. Acetylcholine (ACh), the ubiquitous transmitter in vertebrates has not been detected, although the membrane bound ACh-receptor and ACh-esterase were demonstrated, recently confirmed by genome analysis (Chapman et al 2010). The ACh-receptor will give fast, short-lasting effects in the target cell (maximally tens of milliseconds range), the neuropeptides tend to evoke longer lasting responses (from seconds up to an hour Pol et al 1996). All these aspects: no endplate, innervation by close apposition 'en passant' and neuropeptides next to small-molecule transmitter vesicles are still found in autonomic nerve endings, as will be detailed below. In this respect, the autonomic nervous system can be considered the evolutionary oldest part of the what makes up the vertebrate nervous system.
1.2.2. Embryonic development; origin of the autonomic nerve cells.
In Hydra, the nerve cells are specializations of the epithelial cells that make up the outer and inner layer. In vertebrates, this is still the case: the central nervous system develops as a longitudinal thickening of the cell layer—a placode—on the dorsal (ectodermal) surface of the embryo, which folds up to form a canal. Just before closing the canal, cells from the crest swarm out to the periphery, where they transform into a wide number of specialized cells under the influence of the appropriate inductor genes (Purves et al 2004, Squire et al 2008). For the present introduction, be it sufficient to name the dorsal root ganglion cells (see figure 1), the sympathetic ganglia, the chromaffin cells of the adrenal medulla and the enteric nervous system.
1.2.3. The smallest functional unit.
In motor control of the somatic nervous system (SNS) we have the concept of the 'motor unit', i.e. one motor nerve fiber and the muscle fibers it innervates. Is a comparable unit discernible in the ANS? This question has come up before (Purves and Wigston 1983, Purves et al 1986) and was reviewed by Jänig in Jänig (2006). The answer immediately shows the peculiarities of the ANS: in most instances there is no well-definable autonomic motor unit, connecting one motor fiber exiting the CNS to activate in an all or none fashion a specified number of autonomic effector units.
In the first place, there is the fact that each autonomic motor fiber from the CNS has to pass its message on to a ganglion cell. This is, on most occasions, not a one on one transmission: often the first, so-called preganglionic fiber will branch into a number of terminal fibers. Even if one or more ganglion cells receive sufficient activation to fire in response to each action potential of the incoming motor command, some surrounding cells may also, subliminally, be activated. The incoming activation not only releases acetylcholine that elicits an immediate, short-lasting response in the next, postganglionic neuron, [► at the same time it may release neuropeptides as well, which have much longer lasting effects. Consequently, more incoming activity, either from other preganglionic neurons or from the same one, on repeated activity, may bring that 'subliminal fringe' (Alonso-deFlorida et al 1991) to fire as well ◄].
This is only what might happen in a 'simple' ganglion, like those found in the sympathetic chain. Many parasympathetic ganglia are more complicated, like for instance the cardiac ones. Their proper name has become 'ganglionated plexus' rather than ganglion, not only because of its intricate anatomy, but also since it is more than a relay station: it has integrating properties of its own (Armour 2004, 2008). In the cardiac autonomic plexus, various cell types have been identified, not just postganglionic relay cells, waiting to be activated by preganglionic activity: these cells are spontaneously active, showing subliminal oscillations, which in frequency and amplitude are modified by incoming parasympathetic and sympathetic nerve activity and circulating hormones, thereby making the likelihood of a response to an incoming parasympathetic command dependent on the instantaneous condition. Moreover, local mechanoreceptors from the heart itself may synapse in the plexus as well, thereby serving the shortest feedback loop possible.
The situation in the sympathetic paravertebral ganglionic chain is a little more complicated than depicted above: one finds their preganglionic motor cells in the spinal cord from the lower cervical to the upper lumbar segments, where motor fibers leave the CNS with the somatic motor fibers, to join with the incoming sensory fibers into the spinal nerve. This passes through the opening between the vertebrae to serve its body segment (and/or its derivatives: e.g. the arms derive from the original lower cervical and upper thoracic dermatomes in the embryonic stage). Immediately after leaving the CNS the autonomic preganglionic fibers branch out from the spinal nerve in a whitish connecting branch to meet the segmental paravertebral ganglion or a prevertebral ganglion (see figure 1). These connecting branches are a shiny whitish, due to the supporting cells that isolate the nerve fibers electrically and improve conduction velocity: The Schwann cells, providing the myelin sheets. Below the last white connecting nerve, the sympathetic chain of ganglia just continues as if nothing has happened, showing a segmental ganglion at the respective lower segments, each ganglion recognizable by the greyish connecting fiber that runs back to the spinal nerve, grey due to the lack of myelin around the postganglionic nerves. Grey fibers from the prevertebral ganglia join the arteries to the various organs that are served from their position at the aorta or, like in the lower abdomen and pelvis, they form a separate autonomic nerve innervating the local organs.
The arrangement as described can only work if the incoming motor neurons branch to higher and lower segments before making a synapse in other segmental ganglia than where they started. In the neck, above the segments where sympathetic motor nerves are found, also 3 sympathetic chain ganglia are observed, that derive their motor neuron input from the lower cervical and upper thoracic segments. The branches from these cervical ganglia reach, among other body parts, the heart, the arm and the head, inside and out (figure 3).

Figure 3. Schematic overview of sympathetic and parasympathetic innervation. The organs have been labeled on the right, on the left only those parts that are particular to the sympathetics. Dotted lines: preganglionic, solid: postganglionic fibers. Note the left/right differences in length in that respect. Picture adapted from Low, Clinical autonomic disorders, 1993, first edition, 2nd chapter by Y Harati on: Anatomy of the spinal and peripheral autonomic nervous system, reproduced by permission.
Download figure:
Standard image High-resolution image[► Just looking at this intricate layout from pre- to postganglionic nerve fibers it is clear that the message from start to finish is not carried one-on-one. Along the road, the message splits up, and undergoes interaction with other ones, both from the 'own' branch -parasympathetic or sympathetic- and from the 'counterpart', and it is possibly modulated by circulating hormones and local tissue factors. As a consequence, autonomic responses are variable and one may see activity 'overflowing' from one set of targets to another one, like heartbeat increase and hand-sweating may go together in a lie detector test. ◄]
In search for the motor unit's target organs the complexity is still not over: most postganglionic fibers do their job in the target organ 'by passing by' rather than finding a well-defined motor endplate. The fibers show 'varicosities' i.e. swellings in their fine terminal branches, where transmitter vesicles are found, both dense-core and bright, carrying acetylcholine if parasympathetic or noradrenaline if sympathetic, with and without neuropeptides respectively. Places of close apposition of the preganglionic varicosities and target cells are found, where the most efficient transmission can take place. Most parts of the fibers run at a rather large distance from their targets, therefore, it is not amazing to find spill-over from these transmitters in the venous blood that flows from a target area where the ANS is active. And finally, if the targets are, for instance, smooth muscle cells, these may pass the activation along to the neighbors by low-resistance intercellular contacts, just as in Hydra. This, however, depends on the specific target organ: in the intestinal wall, many close contacts between smooth muscle cells exist, in organs that are supposed to act fast and coordinated, this is much less the case, as in the pupillary muscle to constrict the pupil or in those of the ejaculatory duct of the male reproductive system.
In short, the search for the smallest functional unit of the ANS taught us a lot about the organization of it, but the whole idea of an autonomic motor unit proved to be fuzzy. [► Maybe that is the better way of looking at it: the peripheral organs have an organization of their own, which is modulated by the ANS, adapted as it were to the benefit of the whole organism. This adaptation of innervation specificity is probably established by the interplay of the peripheral tissue and the developing ANS during the embryonic stage (Purves et al 1988). ◄]
1.3. Parallel innervation by the sympathetic and parasympathetic system
One of the first facts about the two opposing branches of the ANS learned from physiology textbooks, is the antagonistic innervation and the balance between the two: more sympathetic activity implies parasympathetic withdrawal and vice versa. It has led to elaborate schemes to quantify the sympathetic/parasympathetic balance, in particular from heart rate variability, but seldom the question is raised, 'is this true', or rather, 'does it exist'? It presupposes two things: (1) a dual (and reciprocal) innervation by both systems and (2) some magic 'balance' at a central level between them that is taking care of this reciprocity.
In his book (Jänig 2006), Jänig is rather outspoken about the first of these issues: the supposed dual innervation is only present in very few target tissues. Most ANS targets are either sympathetic or parasympathetic, only a few receive nerve fibers from both parts of the ANS. However, if considered less strictly, on the level of the organ, reciprocal innervation does exist, although it is useful to keep in mind that the opposing functions may be executed by separate cells. In table 1 is a list of organs with their innervation, sympathetic and/or parasympathetic and the effect that each system has on that organ.
Table 1. Autonomic nervous system functions (see Robertson et al 2012).
| Organ | Sympathetic activation | Parasympathetic activation |
|---|---|---|
| Eye | ||
| Pupil | Dilation (dilator muscle) | Constriction (sphincter muscle) |
| Ciliary muscle | Relax (far vision) | Constrict (near vision) |
| Lacrimal gland | Slight secretion | Secretion |
| Salivary glands | Slight secretion | Secretion |
| Heart | ||
| Rate (sinus node) | Increase | Decrease |
| Contractility (muscle) | Increase | Decreased or none |
| Lungs | Bronchodilation | Bronchoconstriction |
| Gastrointestinal tract | Decreased motility | Increased motility |
| Kidney | Antidiuresis (decreased output) | None |
| Bladder | ||
| Detrusor muscle | Relaxation | Contraction |
| Sphincter | Contraction | Relaxation |
| Penis | Ejaculation | Erection |
| Clitoris, labia minora | None | Swelling/erection |
| Nipples | None | Erection |
| Sweat glands | Secretion | Palmar sweating |
| Piloerector muscles | Contraction | None |
| Blood vessels | ||
| Large arteries | Constriction | None |
| Arterioles | Constriction | None |
| Precapillary sphincters | Constriction | None |
| Venules | Constriction | None |
| Large veins | Constriction, volume mobilization | None |
| Coronary arteries | Dilatation | None |
| Muscle | ||
| Arterioles | Dilatation | None |
| Metabolism | Increased | None |
| Muscle spindles | Decreased sensitivity | None |
| Liver | Glycolysis, glucose mobilization | Glycogen synthesis |
| Fat tissue | Lipolysis | None |
| Immune system | Suppressed | Activated |
This list is by no means exhaustive, it is just showing that the ANS has a grip on all body functions. To fulfill this task, the ganglion cells must produce rather specific output patterns, differing per target tissue (vascular smooth muscle requiring command strings which are very different from the one going to the vas deferens). These patterns, somehow, must be run as a subroutine in the interplay between descending supraspinal command neurons in the CNS, autonomic motor neurons, incoming afferent signals from the target tissues and adjacent tissues, interneurons and ganglion cells. [► It is not readily conceivable that this 'autonomic subroutine' is, so to speak 'prewired', in the genetic code and will be implemented once the hardware is fully developed during embryonic and fetal development. I tend to favor the view that these subroutines of the ANS come into being during development, in a tight learning symphony between target tissue, ingrowing efferent postganglionic nerve and afferent feedback from the tissues. ◄] In that way the ANS behaves in the same way as the motor part of the CNS: movement is a learning process, which begins in the womb, and may end only years later, like when an athlete has mastered his run up and long jump to expert level.
In conclusion to this heading: it is more precise to speak of the parallel control by sympathetics and parasympathetics of a particular organ than to stress their antagonistic actions. Moreover, the individual control systems per organ are probably not organized 'top down' but 'bottom up'—the interaction of the target tissue with the incoming nervous outgrowth in the embryonic (and later) development molds the control system to its specific function.
1.4. Function of ganglions and ganglionic plexuses; the enteric nervous system, how many 'little brains' do we have?
The 'reason' implied in the autonomic regulations defies the higher reason from the CNS. Particularly the intestinal tract is self-regulated, after the transition from striated to smooth muscle at the entrance of the pharynx. This to such an extent and with such an elaborate onboard nervous system that the enteric nervous system (ENS) got nicknamed 'the little brain': it has some 200–600 million neurons (Jänig 2006), loosely organized in two layers. By analogy, since more and more autonomic ganglionic plexuses were shown to have a complex organization as well, sometimes they are also thought of as 'little brains' (Armour 2008). What these plexuses2 have in common is that they constitute an organized network of parasympathetic preganglionic fibers and—postganglionic neurons, incoming postganglionic sympathetic nerve fibers, synapsing side branches of visceral sensory axons and integrating neurons, or 'local circuit neurons' as Armour calls them in the cardiac plexus.
1.5. Transmitters and co-transmitters, pre- and post-ganglionic
The textbook organization of the ANS and its transmitters may be tough to memorize at the first encounter in the science curriculum. When reality kicks in, it gets even worse. As already noted for Hydra, the transmitters for its supposed synapses between nerve and muscle are a mixed bag: fast acting small-molecule responses, combined with longer duration neuropeptide ones. In the vertebrate ANS it is the same story, but since there are at least 2 synapses involved with different properties, it is even more complicated. The neuropeptides involved are different at the pre- and post-ganglionic sites and different for the parasympathetic and sympathetic divisions, and even different in different tissues. This relates to what I described above as the 'local interaction' between the ingrowing nerves and the tissue at hand. All in all, it makes for a complex interaction between the pre- and postganglionic neurons and then from the postganglionic fiber to the effector organ. The 'simple scheme' tells that in the ganglion ACh is the general transmitter, both for sympathetic and parasympathetic ganglia. The postganglionic sympathetic transmitter is norepinephrine (NE), except for the adrenal medulla which secretes epinephrine (E) and the sweat glands where ACh is the transmitter again, however hitting a different receptor than the ACh-receptor in the ganglia. The postganglionic parasympathetic transmitter is ACh. The ganglionic ACh-receptor is sensitive to nicotine (N-type), the postganglionic ACh-receptor is sensitive to muscarine (M-type). These 2 compounds can be considered as pharmacological tools which help in typifying the receptor's molecular structure. Table 2(A) describes the classical scheme, table 2(B) enumerates the co-transmitters, neuropeptides and other non-adrenergic, non-cholinergic transmitters (NANC) which are secreted in specific tissues instead of, or in parallel to the 'classical' transmitters of table 2(A). The complex of transmitters that go together for a specific function goes under the name of 'neurochemical coding' (Elfvin et al 1993). Remarkably, the multitude of neuropeptides is known for its widespread hormonal effects when administered systemically, however, their 'true' effect is still largely unknown. [► The most probable physiological effects are those that are due to so-called paracrine excretion during nerve activity. This may not occur at every one action potential, but has been shown to, definitely occur at higher frequencies of nerve activity (Purves et al 2004). Transmitter overflow might, occasionally, provoke some systemic effects as well. The true function of the neuropeptide co-transmitters is still very much debated ◄]
Table 2. ANS transmitters and putative co-transmitters (see Robertson et al 2012).
| Organ | Sympathetic system | Parasympathetic | ||
|---|---|---|---|---|
| 2A classical transmitters | Transmitter | Receptor | Transmitter | Receptor |
| Ganglia and adrenal medulla | ACh | N | ACh | N |
| Sympathetic effectors | NE | α or β | ||
| Except sweat glands | ACh | M | ||
| Parasympathetic effectors | ACh | M | ||
| 2B putative co-transmitters | ||||
Sympathetic neurons from
|
||||
|
||||
| Chromaffin cells of adrenal medulla | E, NE, Enk, NPY | |||
| Parasympathetic neurons from major ganglia | ACh, VIP, SP, NPY, NO | |||
For the acetylcholine receptors: M, muscarinic; N, nicotinic; 5-HT, 5-hydroxyltryptamine; ACh, acetylcholine; CCK, cholecystokinin; DYN, dynorphin A (DYN 1-8, or DYN 1-17); E, epinephrine; Enk, enkephalin; NE, norepinephrine; NO, nitric oxide; NPY, neuropeptide Y; SP, substance P; VIP, vasoactive intestinal polypeptide.
2. Innervation of some specific organ systems, their reaction to sympathetic- versus parasympathetic activity
In table 1 the autonomic innervation of a number of organs has been pointed out, with the specific effects of the sympathetic or parasympathetic activity. In view of the scope of the journal a logical question is, how to measure those effects. Some of these are straightforward, for instance clinical chemical measurement in the blood will measure changed glucose levels or free fatty acids in response to sympathetic activation. I will give a few examples of specific organ activity and its measurement, all the while showing some peculiarities of the autonomic nervous system.
2.1. Pupil diameter and accommodation to a nearby object; autonomous oscillations of pupil size
The objective of the system is to adapt the pupil size to the amount of light that may be admitted to the retina for proper image formation. For that purpose, two, each other opposing, smooth muscles are available in the opaque iris: the parasympathetic sphincter pupillae and the sympathetic dilator pupillae. So, in short, we have a feedback system at hand: not the rods and cones of the retina, but the intrinsically photosensitive retinal ganglion cells (Do and Yau 2010, McDougal and Gamlin 2015), which send out an integrative signal, conveying the illumination strength to the midbrain, and 2 effectors, of which the sphincter is the fastest with a motor nucleus close by (probably the Edinger-Westphal nucleus in the oculomotor complex of the midbrain, but see Sun and May (2014) for diverging opinions on the exact pathways in different species).
The illumination signal is independent of a connection to the optical cortex; subjective awareness of the signal is not required to elicit a pupillary response to a bright light. Not only is the pupillary constriction a parasympathetic function, regulated from this site, so is the innervation of the ciliary muscle, that induces bulging of the lens inside the eye for close by vision. Being part of the oculomotor complex, the whole system of accommodation and pupillary constriction is also coupled to convergence of the eyeballs. This whole set, pupillary constriction (also called miosis), accommodation of the lens and convergence goes together under the name of accommodation reflex. Why it is necessary to combine pupillary constriction in this 'near response triad' is not clear.
The way to parasympathetic pupillary constriction may be more or less straightforward, the one to sympathetic pupillary dilatation is less obvious. The 'classical' motor neurons are located in the lower cervical, upper thoracic spinal segments (all sympathetic motor neurons are in the spinal cord). Additionally, the parasympathetic outflow may be diminished at the midbrain level via an adrenergic pathway (Thompson, chapter 41, Primer, Robertson). Question for the pupillary dilatation is, what is the command input? Is absence of light a stimulus for pupillary dilatation or is it the consequence of relaxation of the sphincter muscle? Is the maximal pupil size in the absence of light set by a 'central sympathetic tone' which can be considered a reflection of a mental state of more or less excitement?
To get a little closer to an understanding of the problem we should look into measurement of pupil size. Thanks to miniaturization and simplification by solid-state techniques it has become rather easy to obtain a continuous curve for the pupil diameter by the use of infrared light emitting diodes and continuous analysis of the reflected image in which the pupil is a dark spot (Larson and Behrends 2015). This equipment enables the clinician to quantify the pupillary light response (PLR) and the pupillary reflex dilation (PRD). The first thing, however, that comes out is the instability of the pupillary signal in the awake test person. Changes in gaze and accommodation lead, as explained above, to concomitant changes in pupil size. On top of that, in particular, when the test subject is drowsy, the pupil diameter shows small oscillations around the respiratory frequency, which have become known as 'the hippus'. This may also be observed under light anesthesia; it disappears when the anesthetic level deepens (experimentally studied in cats (Borgdorff 1975), but applicable to humans as well, as reviewed in Heaver (2011) and Onorati et al (2013). Borgdorff (1975) conclusively showed that the hippus reacts to baroreceptor input and is only dependent on parasympathetic activity. [► The sympathetic contribution to pupil size may, therefore, be considered a reflection of central sympathetic state, more than anything else. ◄]
In view of its properties as a feedback system, light stimulation by a small spot light source on the edge of the pupil, a sustained oscillation may be expected. This has, indeed, been observed, and in a good linear approximation the system can be described as a third order integrating system with a pure time delay (Khoo 2000b, p 46; τ = 0.1 s, D = 0.18 s). In conclusion, one may say that (in health) the pupillary diameter is the reflection of a feedback system, regulated via the parasympathetic efferent channel and an adaptation to the internal and external environment (flight, fright, fight) under sympathetic control.
2.2. Heart, contractility, heart rate and heart rate variability
At the gastrula stage in embryonic development a new germ layer is formed between endo- and ectoderm: the mesoderm. Very early along the midline two mesoderm-derived vascular tubes appear that fuse in the midline and a primordial heart (still without valves) starts to beat. The cells of this tube all have pacemaking properties, but the fastest pacemaking takes place at the inlet of the developing heart, conducting the stimulus along the tube in a peristaltic wave. Under the influence of the surrounding structures (notably the primordial vertebral column) and helped by cells that stem from the neural crest, this tube heart develops into the final four chambered mammalian and avian heart serving a double circulation: pulmonary and systemic. The embryonic history is still worth knowing: be it alone to understand where the heartbeat is initiated, i.e. in the area of the original entrance to the tube heart: the sinus venosus, the place where the venous blood comes together.
In essence any adult myocardial cell can still act as a pacemaker cell, if the circumstances allow it. Notably some loss of resting potential due to hypoxia or changed plasma levels of important ions like sodium, potassium or calcium, may bring about spontaneous depolarization. Luckily, most of these cells will be quiet, though, and only the sinoatrial (remnant of the sinus venosus, the inlet of the primordial heart) and atrioventricular nodes will show spontaneous depolarization that is able to initiate a heartbeat at physiological heart rates. The cells of the ventricular conduction system may do so as well, however, at very low rates, barely sufficient for survival.
As shown in table 1, the heart is under the influence of both the sympathetic and parasympathetic nervous system, being served from the cardiac plexuses, mainly located around the atria. These sites may be considered 'integration sites' where efferent autonomic and afferent cardiac (contraction, interstitial condition) come together to influence heart rate and cardiac performance.
As was the case for the pupil of the eye, here too the parasympathetic influence is the fastest and most dominant regulator of (mainly) heart rate. When left to itself the sinoatrial node produces a rate of around 100 bpm, the so-called intrinsic heart rate, which in vivo may be observed after combined muscarinic (parasympathetic) and β-adrenergic blockade; its value can be approximated by 110 − 0.6*age (Opthof 2000). This shows that during most of the day the heart is mainly under parasympathetic control, when heart rate remains between 60 and 100 bpm. Only during exercise it will get higher, to max out at a value of around (208 − 0.7 * age) (Tanaka et al 2001) (Note: this is an updated version of the weakly based older formula of (220 − age)). Heart rates at maximal exercise will scatter widely anyway. The old formula tended to underestimate the HR to be obtained by subjects over 40.). In health, the most important determinants of heart rate variability are blood pressure via the baroreflex and respiration. Blood pressure control is treated in more detail in the next section ('Blood vessels'), be it sufficient here to point out the ubiquitous presence of pulse pressure related activity in the nervous system. As shown in figure 4 the pulse wave induces a burst of incoming activity via the baroreceptive afferent nerves which are connected to stretch receptors in the walls of a number of large arteries. This provokes an almost immediate response in the efferent cardiac vagal fibers to dampen heart rate. After all, the pressure receptors are located in remnants of the original branchial arteries, dating back to the fish-state in evolution. Too high a pressure in the gills cannot be tolerated by the delicate microvessels which subserve the oxygen uptake from the surrounding water. Mean resting heart rate is the product of this repeated vagal bombardment (and of background sympathetic and hormonal activity). Alterations in the incoming traffic, due to changes in pulse—and/or mean pressure, will be mirrored by changed vagal efferent activity with heart rate effects within the same beat at resting rates below ~75 bpm; at higher resting rates the next beat will be changed. (Pickering and Davies 1973, Karemaker and Borst 1980), leading to less brisk effects and apparent slowing of the reflex time from baroreceptors to heart rate (Karemaker 2015).
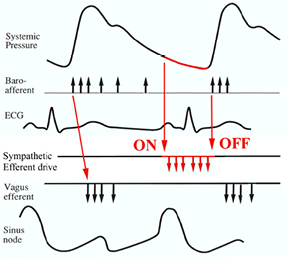
Figure 4. Schematic time relations of baroreceptor afferent activity and feedback via sympathetic and vagus nerve efferents to the sinus node. The cardiac cycle starts at the sinus node depolarization (lowest trace). This initiates the usual sequence of the ECG. The pulse pressure (top trace) induces baroreceptor afferent activity, which in turn triggers vagus nerve activity. Sympathetic activity is unleashed by the absence of baroreceptor afferent activity. The vagus nerve volleys may induce cardiac slowing by decreased diastolic depolarization of the sinus node or a complete reset to its starting level, as indicated. Figure after Karemaker and Wesseling (2008), by permission.
Download figure:
Standard image High-resolution imageApart from its effect on heart rate, the parasympathetics will decrease contractility, much so in the atrium, but also, slightly, in the ventricles (Kent et al 1974, Lewis et al 2001); this effect goes beyond the effect of just the decreased heart rate. Moreover, vagal activity will decrease the conduction speed through the AV-node, but also in the atria and ventricles. The latter may be beneficial in case of reentrant tachycardia's, be they atrial or ventricular or a combination as in the Wolff–Parkinson–White syndrome where an extra conducting bundle of Kent exists between atrium and ventricles. Vagal stimulation as for instance induced by carotid sinus massage may stop the reentry due to the decreased speed of conduction.
The sympathetics innervate all of the heart: the atria, with the sinoatrial and atrioventricular nodes, and the ventricles. The β-adrenergic receptors (of both the β1- and β2-type) are activated by circulating adrenaline (β1 and β2) and locally liberated noradrenaline (β1) from postganglionic nerve endings. Sympathetic activation increases heart rate and contractile force at the expense of increased metabolic demands; therefore, this activation is deleterious under conditions of a compromised coronary circulation.
The study of heart rate variability (HRV) has witnessed an exponential growth in publication numbers since the seminal papers from the groups of B. McA. Sayers at Imperial College, London (Hyndman et al 1971, Sayers 1971, Kitney and Rompelman 1980) and, later, R J Cohen at Harvard-MIT (Akselrod et al 1981, Akselrod et al 1985). The initial period was dominated by linear analysis, fast Fourier transform and autoregressive methods to obtain in particular well-defined numbers for low- and high-frequency oscillations (LF and HF) in heart rate. HF (around 0.25 Hz) being related to respiration via the parasympathetics and LF (around 0.1 Hz) to a sympathetic Eigen-oscillation of the baroreflex to blood pressure and heart rate system (DeBoer et al 1987). This sparked the idea of the LF/HF ratio that would point out the central nervous balance between the sympathetic (stress) and parasympathetic (relaxation) systems. This idea was pushed strongly by Malliani and his group in Milano (Pagani et al 1984, 1985, Baselli et al 1986, Lombardi et al 1987), which helped opening up the area of HRV-analysis for basic research and clinical applications. The idea was not undisputed, though, most fiercely objected over the years, among others, by Taylor et al (1998) and Bertisch and Taylor (2011) as to the relation between LF and sympathetic activity. Recently (Draghici and Taylor 2016) gave an updated review of these objections and, among other, against the use of LF/HF, in particular against the view that HF might be a reliable measure of parasympathetic outflow; I will come back to this issue below.
The fact that HRV was shown to correlate strongly with outcome after myocardial infarction (Kleiger et al 1987) sparked the interest of the cardiologists at large in HRV for practical purposes. It had become clear that HRV is mainly determined by the parasympathetic influence on the sinus node: this is able to cause fast beat-to-beat jumps in sinus rhythm (specific pathologies of the pacemaking process excepted). Many of the developed tools to put a number to HRV have become part and parcel of the diagnostic package, in particular when judging 24 h (ambulatory) Holter-recordings. A short list of the numbers that are in use and how these are calculated can be found in table 3. However, until now HRV is still not more than at best an adjunct diagnostic tool in most cardiology clinics. In particular, since it was established that it was really an epi-phenomenon and not something that could be used to improve the patient's condition by, pharmacologically, boosting HRV. This has been tried, in vain, by low-dose atropine which is known to increase vagal outflow from the CNS to the heart rather than blocking it at the sinus node muscarinic receptor, as a high dose would do. Scopolamine dermal patches, used for the same purpose as low-dose atropine, led to lower HR and increased HRV, but it did not improve exercise performance or the severity of ventricular arrhythmias (Porter et al 1990, Vybiral et al 1993, Casadei et al 1996a).
Table 3. Heart rate variability measures in frequent use (see Billman 2011).
| Time domain | Units | Description |
|---|---|---|
| SDNN | ms | Standard deviation of all Normal-to-Normal beat intervals; considered a vagal measure of short-term variability |
| RMSSD | ms | Square root of the mean squared differences between N–N intervals; considered a measure of short-term (vagal) variability |
| SDANN | ms | SD of period-averaged N–N intervals (usually 5 min averages); computed over (24) hours, considered a measure of long-term variability |
| pNN50 | % | Percentage of all adjacent N–N intervals that differ by more than 50 ms |
| Frequency domain | ||
| Total power | ms2 | Area under the entire spectral curve; total variance |
| ULF | ms2 | Ultra-low frequency: power in the band below 0.003 Hz; requires the longest recordings to reliably be measured (>10 min at least) |
| VLF | ms2 | Very Low Frequency: power from 0.003 to 0.04 Hz |
| LF | ms2 | Low Frequency power from 0.04 to 0.15 Hz; considered a measure of sympathetic activity in blood pressure control |
| HF | ms2 | High Frequency power from 0.15 to 0.4 Hz; considered respiration-coupled vagal heart rate modulation |
| LF/HF | Ratio of LF to HF power. May be computed after normalization of LF and HF to a total power just represented by (LF + HF) or directly | |
An issue that warrants a separate discussion is the exact physiological background of the above-mentioned HF variability or respiratory sinus arrhythmia (RSA) in heart rate. One thing is certain: it represents a respiration-related modulation in parasympathetic outflow to the sinus node, but how does it come about and is all of the parasympathetic outflow subject to this modulation? These questions have found an intricate answer in recent experiments from McAllen's group (Farmer et al 2016). They elegantly demonstrated the participation of at least three separate centers in the brainstem of the rat in the maintenance of tonic and respiratory modulated parasympathetic activity. Part of it is due to direct interaction of respiratory centers and vagal motor neurons, another part to the NTS (cf. fig.6), not baro- or chemoreflex dependent, and part is of, as yet, untraced origin. In the awake preparation that was used by these authors, blood pressure was low and more or less constant, therefore any baroreflex modulation by respiration-induced blood pressure waves was absent, actually a fourth possible origin of modulated vagal outflow—be it caused by respiratory or slower blood pressure waves (like sympathetically mediated 10 s waves). These findings throw a new light on many earlier unresolved issues about the origin of RSA and its relation to cardiac vagal tone: the corollary that RSA would be a good measure of vagal activity is, clearly, untenable; RSA cannot completely be explained by baroreflex feedback either, and the amount of HF-activity in the spectrum of HRV does certainly not put a number to a central 'resting state' condition. Is this the end of the LF/HF story? Not quite: for instance, when someone is tilted gradually, from relaxed supine to almost upright (supported) standing on a tilt table, a concomitant change from little LF and much HF to, finally, much LF and little HF can be observed. Here, the LF/HF ratio does follow the transition from a parasympathetically dominated rest to a condition where the sympathetic system must be prominently active to maintain upright blood pressure (Cooke et al 1999). This shows that, within limits, frequency analysis of HRV can give a description of a state change in the ANS. Of course, when the system is approaching saturation levels, like in heavy exercise (Casadei et al 1996b), where heart rate is reaching extreme values, this analysis will fail: in order for HRV to give information there should be variability.
The linear techniques of time- and frequency domain analysis from the first period of HRV-analysis, have been complemented with non-linear techniques, which, generally speaking, look at the behavior of the sinus node oscillator in a multidimensional phase space, where it turns its rounds in more or less complex trajectories, possibly being thrown of course by a multitude of factors inside and outside the heart (or even the subject). One way to find order in this more or less chaotic behavior is to look for repetition of interval sequences within a set tolerance and attach a predictability number to that series, derived from information theory (i.e. entropy Pincus 1991, Richman and Moorman 2000). A historical overview of the various applied analysis techniques has been given by Billman (2011); this publication is part of an e-book on the subject of HRV and clinical applications (Trimmel et al 2015). However, these approaches have their opponents as well: for one it has been argued that most of the resulting numbers can be supplanted by just stating the heart rate (Stauss 2014)—higher heart rates implying less variability in whichever way it was measured. At least should the measures as mentioned in table 3 be corrected for heart rate to make sensible clinical decisions (Monfredi et al 2014). Another objection is the lack of repeatability of the measurements, even in healthy subjects: the 24 h hrv spectrum on a log–log scale is fairly linear, apart from some bumps around 0.1 Hz and the respiratory frequency, which reminded of a fractal process since a heart rate recording at various time scales (from seconds to minutes to hours) gives almost the same impression of 'jumpiness' (Peng et al 1995, Makikallio et al 1998). Logically, the negative slope of the log–log spectrum would, then, count as a measure to describe this process and, maybe, show pathology by its shifting outside a 'normal' range. Tan et al (2009) showed the fallacy of this approach by repeated measurements: the fractal scaling exponent could considerably change from session to session, thus the authors concluded that this could not be used as diagnostic tool.
Until now one of the few clinically applied uses of non-linear HRV-analysis is in the proprietary monitoring program by Moorman et al which is intended for the neonatal intensive care unit (NICU) to find early indications of sepsis (Fairchild et al 2013). Figure 5 is taken from their 2011 publication (Moorman et al 2011) which led up to the development of the NICU monitoring system: it shows clearly how the normal HRV dies out and more and more HR-decelerations occur in the wake of a developing sepsis. Obviously, these decelerations are bursts of parasympathetic activity (interruptions of sympathetic activity would be much slower and less 'pointy' in aspect). I will come back to the connection between the immune system and the ANS later. The disappearance of the fast, beat-to-beat variability can, in terms of the multidimensional phase space, be regarded as a loss of more and more dimensions, i.e. factors that impinge on the heart rate driving neural centers, in particular that of the vagus nerve. Other conditions can have this same effect, like anesthesia (Scheffer et al 1993) or coma (Machado-Ferrer et al 2013).
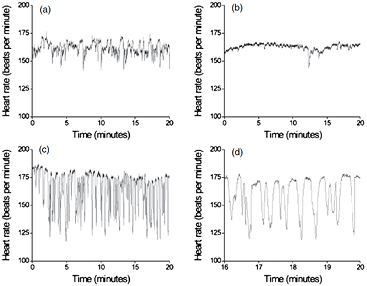
Figure 5. Time series recording in one neonate. (a) Normal HRV; (b) reduced variability; (c) storm of heart period decelerations [► probably of vagal origin ◄]; (d) detail from recording in (c). Recordings (b)–(d) were made hours before (other) clinical signs of sepsis were present. Illustration from Moorman et al (2011), by permission.
Download figure:
Standard image High-resolution image2.3. Blood vessels and blood pressure control
2.3.1. Flow and pressure at a global level.
To have effective nutrient flow through the microvessels a minimal pressure higher than the plasma colloid osmotic pressure (around 28 mmHg) is required. Taking into account pressure losses in the conduit arteries and draining venules and veins the driving arterial pressure should be at least 50 mmHg. These numbers go up if one looks at the kidney which has higher requirements for driving pressure in view of the ultrafiltration process in the glomeruli. And finally, if the head is held above the heart, a hydrostatic column should be added to this, more for the giraffe than for the average human, bringing the lowest number for a 'normal' upright mean arterial pressure (at heart level) to some 75 mmHg. The ANS is called in to maintain this pressure. Ohm's law for the circulation will tell that Pressure = Cardiac Output * Resistance, therefore, vascular resistance, together with heart rate and stroke volume are the determinants to control blood pressure. Blood vessels are richly innervated by the sympathetic nervous system, apart from a few specialized vessels (e.g. in erectile tissues) the parasympathetics have generally no business with them. This leaves it to the sympathetic system to regulate pressure and distribute flow at a global level, in close cooperation with local mechanisms for the fine-tuning of supply and demand.
The active component of blood vessels that may modulate their diameter is the vascular smooth muscle cell (VMC). Depending on their place in the vascular tree, the vessel walls have various amounts of VMC's, ranging from almost none in the mainly elastic wall of the aorta to very little elastin and almost only VMC's and endothelial lining cells in the arterioles and small diameter veins (Burton 1972). The presence of α-or β-receptors in their various subtypes is dependent on the specific tissue where one is looking and the same holds for the purinergic receptors (Robertson et al 2012). This makes sense if one realizes that sympathetic activation may lead to contraction of skin vasculature as protection against the cold, but also to dilatation of the vessels in the active muscles. The latter has been a longstanding issue in exercise research: is there a mechanism for active dilatation of the vessels that is activated at the beginning of exercise, by some form of 'central command' in parallel to the activation of the muscles? The mechanisms of adaptation of muscle flow to demand have recently been reviewed (Joyner and Casey 2015), where the authors make the case that vasodilatation in exercise is, in the first place, due to a decreased sympathetic vasoconstrictor tone, which under resting conditions will restrict muscle flow. On longer duration of exercise local mechanisms take over, for instance by the flow-mediated vasodilatation which is a function of the endothelium under increased flow stress, when it is producing vasodilating factors like NO as recently reviewed by Gutterman et al (2016). Still this is not the full story, for instance in the defense response, known to cause immediate vasodilatation (Roddie 1977), other mechanisms are probably involved, like cholinergic or purinergic sympathetic activity (Joyner and Casey 2015).
Central control mechanisms for blood pressure require afferent signals to act upon: these are found in the 'reflexogenic' areas of the circulation, of which there are many (Heymans and Neil 1958), but here restricted to the so-called baroreceptive areas, places where the arterial walls are sensitive to stretch. This stretch is provided by the arterial pulse wave, so the incoming signal is 'pulsatile' as well, as indicated in figure 4, sensitive to the height of the pulse wave and to the local stiffness of the wall where the sensors are attached to.
Not only the driving pressure but also its effect, i.e. how well the organ perfusion is able to maintain homeostasis, is turned into incoming signals: when oxygen delivery or waste removal fall short, metaboreflex afferents are activated (Nobrega et al 2014). Figure 6 from Karemaker and Wesseling (2008) represents our view on how the CNS handles the various incoming signals and transforms these into the autonomic output signals as we may observe them, be it from their effect on HR (parasympathetics mainly) or in the sympathetic efferent signals that can be recorded by transcutaneous microelectrodes in peripheral nerves. Many neurologists have learned this technique and how to distinguish the signals from these fibers from other efferent or afferent traffic thanks to the work of a Swedish group of neurologists (Hagbarth and Vallbo 1968, Delius et al 1972, Hagbarth et al 1972), in recent years mostly represented by Wallin (Joyner et al 2016). Nerve recording has become the gold standard of measurement for sympathetic activity, in particular so for muscle sympathetic nerve activity (MSNA) which was shown to be related to blood pressure control (see figure 4). Skin sympathetic nerve activity (SSNA) is related to temperature and emotion, and generally will not come in arterial pulse-related bursts. One issue of figure 6 should be highlighted, i.e. the box 'max sympathetic outflow', representing a supposed 'central sympathetic excitation' which is mainly set by brain centers higher-up in the hierarchy, psychological factors, sleep/wake cycle, to mention but a few. Furthermore, the chemoreflex afferents (involved in respiratory control) and metaboreflex afferents may, at the level of the medulla oblongata, contribute to this central sympathetic arousal (Dempsey et al 2002).
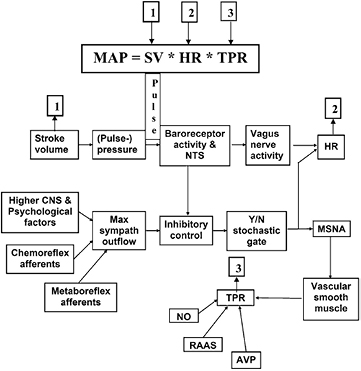
Figure 6. Schematic of baroreflex functioning. The afferent signal (baroreceptor activity, traveling to the NTS = nucleus tractus solitarii in the medulla oblongata) originates as consequence, mainly, of the pulse pressure at the receptors (i.e. stroke volume-related). It translates more or less directly into efferent signals over the vagus nerve to lower heart rate (HR). At the same time, it centrally inhibits sympathetic outflow that after the inhibitory control has to pass some stochastic gates (Kienbaum et al 2001) before being translated into efferent muscle sympathetic nerve activity (MSNA). The latter stimulates vascular smooth muscle to contraction, thus increasing total peripheral resistance (TPR). This, by itself, is under influences like locally produced NO (nitric oxide), kidney- and adrenal-involvement by the renin-angiotensin aldosteron system (RAAS) and by arginine vasopressin (AVP) from the hypophysis. The entities 1, 2 and 3, together, determine mean arterial pressure (MAP) as product of cardiac output and total peripheral resistance. MAP and pulse pressure, together, are determinants for baroreceptor activity, thus closing the feedback loop. Illustration from Karemaker and Wesseling (2008), by permission.
Download figure:
Standard image High-resolution imageThis view may lead to unexpected consequences: Physiology teaches that the sympathetic system will be activated when blood pressure drops. And indeed, this occurs when the subject is in a steady state condition: in particular in the standing position, when (diastolic) BP drops a little, the sympathetics will be activated, resulting in more MSNA (figure 7). Then the resulting increased BP will shut off further MSNA, BP drops again a little, etc (This is the generally accepted explanation for the observed 10 s rhythm in BP as can be shown by a simple computational model DeBoer et al 1987.). However, in the operating room, when anesthesia has crippled the central sympathetic drive, BP will not automatically be restored, since there is nothing to be unleashed when BP drops thereby lifting the restraint due to incoming baroreceptor afferent traffic.
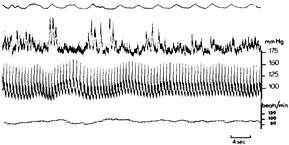
Figure 7. Relation between intra-arterial blood pressure and muscle sympathetic nerve activity (MSNA) recorded in the median nerve. Test subject supine, slightly head up tilted. Upper tracing: Respiratory movements (inspiration upwards); second tracing: Integrated MSNA signal; third tracing: intra-arterial blood pressure (BP); fourth tracing: heart rate (HR). It is obvious that MSNA increases when blood pressure decreases, followed by restoration of BP and increases in HR. Picture adapted from Delius et al (1972), by permission.
Download figure:
Standard image High-resolution imageOne notable exception to the general descriptions of this section is found in the cerebral circulation. It maintains a high degree of 'autoregulation', i.e. the tendency to keep flow more or less constant in the face of changing pressures. It is better able to do this for slow changes than for fast: if it would behave as a perfect differentiator, one would expect to find a phase advance of 90° over the whole frequency spectrum, however in healthy individuals this number is already down to around 60° at a frequency of 0.1 Hz approaching 0° at the usual respiratory frequency of 0.25 Hz, it behaves thus as a differentiator with integrating properties. We owe this insight into the dynamic aspects of autoregulation to the transcranial Doppler devices which enable measurement of cerebral blood flow velocity on a beat-by-beat basis. Together with continuous finger blood pressure (Finapres, and its successors, the original invention of Jan Peñaz and Karel Wesseling, Imholz et al 1998) non-invasive bedside detection of derailment of dynamic autoregulation has become a possibility (e.g. Immink et al 2005). How to treat these signals and compute standardized transfer functions from pressure to flow has been the subject of a recent 'white paper' by an international collaboration on this issue (Claassen et al 2016). However, the underlying physiology of the phenomenon is all but resolved. It is evident, that we are looking at a property of the larger vessels (the microcirculatory aspects will be treated below). These vessels are richly innervated by sympathetic nerves and, in particular the vessels that serve the posterior part of the brain, by parasympathetic nerves (Roloff et al 2016). Still, the function of the innervation is obscure: if someone stands up from supine, systemic vascular resistance will increase by widespread sympathetically induced vasoconstriction. However, if the cerebral vessels would participate as well, brain blood flow would be even more compromised than it already was by the hydrostatic effects on the perfusion pressure. [► This bypasses the question of a possible 'siphon-effect' where the venous downhill flow would support the arterial uphill flow; since the brain microvasculature behaves, technically, as a 'baffle' separating the arterial from the venous side (Gisolf et al 2005) ◄]
2.3.2. Local vascular mechanisms.
The microvessels 'know' much better than the CNS what the local tissue needs. Therefore, much of the regulation is local, in reaction to the surrounding milieu (e.g. hypoxia, CO2 and lactate build-up due to insufficient flow). The adaptation takes place, partly by a direct action on the VMC, partly by locally produced dilating or constricting factors, mainly produced by the endothelial cells. The best known locally acting 'transmitter' is, of course, NO, nitric oxide, that originally became known as a rather volatile EDRF (endothelial-derived relaxing factor). Although the endothelium may produce NO, it is not the only source: it is also produced by the innervating neurons (Chowdhary and Townend 2001). In fact, there is an intricate balance between locally produced vasodilating and vasoconstricting factors and the activity of the sympathetic neurons and circulating hormones. Figure 8 gives an schematic of these influences, a figure taken from the recent book by Jänig (2006), who himself credits Folkow and Neil (in their 1971 book, Circulation (New York: Oxford University Press)) as the originators of this view on the influences on tissue flow.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 8. Multiple control of blood vessels: left: remote; right: local control. Remote are the sympathetic vasoconstrictor neurons; vasodilator neurons (in some tissues only) and blood borne influences like adrenaline, angiotensin II and vasopressin (e.g. during blood loss). Local control as by spontaneous myogenic activity, stretch-induced activity, endothelium induced vasodilatation (e.g. by NO under shear stress due to increased flow). Vasodilatation due to mast cell and other cells like polymorphonuclear leukocytes (PMNL) activity during inflammation. Vasodilatation by peptides released by small diameter afferent nerve fibers. Illustration from Jänig (2006), by permission.
Download figure:
Standard image High-resolution image{kind=link}
The brain microvessels, are exceptional in their structure. One group of the many cells that support the neurons in their function (the astrocytes, part of the so-called glia-cells), have extensions which completely encapsulate the microvessels. Thus, they help form a blood-brain barrier, which shields the neurons from many of the circulating compounds: only those that can 'make it on their own' by being well soluble in the fatty membranes that make up the barrier, or those that are actively taken up will make it into the extravascular space (Bradbury 1985, Lajoie and Shusta 2015). In reaction to neuronal activity, the arterioles serving the particular area will dilate, again under the influence of locally active substances. This dilatation is not implied in the dilatation at the macro-level, as described above under autoregulation, even though some of the same agents may be involved, like NO.
One peculiarity of local flow mechanisms needs to be mentioned here: the rhythmic vasomotion, i.e. contractions of microvessels, loosely influenced by nervous activity, which can be propagated to adjacent vessels and, thus, spread out over larger distances. These rhythms are unrelated to the cardiac or respiratory cycle, and may appear regular at times, and chaotic at other moments. In a recent review on the subject it is argued that vasomotion is actually a network action, where the 'wisdom of the network' (jmk) initiates local dilatations and constrictions to optimize perfusion of the ensemble (Pradhan and Chakravarthy 2011). A complete description of the local interactions of the many processes is outside the scope of this introduction. The interested reader is referred to Aalkjær et al (2011) for a recent update.
2.4. Lungs and respiration
2.4.1. Respiratory rate variability.
Forget everything about required perfusion pressure when it comes to pulmonary flow: here we have a low-pressure system where the intent of the flow is not to be nutrient to the tissues but to have as much of the passing cardiac output saturated with oxygen and get rid of the CO2. The nutrient flow for the lungs is derived from the systemic circulation by way of the tracheal and bronchial arteries. The action of the ANS on the lungs must be viewed in this same scheme: is cardiac output (throughput) or the lung itself at stake?
The lungs receive their autonomic innervation both from the sympathetic and the parasympathetic (vagus) side. As indicated in table 1, the best known influence of these is on the airways; this does not mean that the vessels are not innervated. It only shows how in medical praxis the observable predominates. Problems due to airway resistance are common and well-known (e.g. in bronchial asthma, or in chronic obstructive pulmonary disease); whether the resistance to pulmonary blood flow is increased, will only very late in the disease process lead to the diagnosis of pulmonary hypertension, since the initial symptoms are non-specific: shortness of breath, fatigue, weakness to name a few (Galiè et al 2009).
During the normal respiratory cycle the sympathetic and parasympathetic efferents are active in counter-phase: the sympathetics fire during inspiration, dilating the airways, the parasympathetics in expiration, [► maybe stiffening them and preventing collapse of the floppy smaller airways inside the thorax ◄], alternatively this activity might be related to glandular activity as suggested by Jänig (2006). Indeed, the activity of the vagus nerve has recently been implicated in postoperative acute respiratory distress syndrome (ARDS) as a factor that induces increased airway resistance (Chen et al 2016).
Respiratory rate and depth are adapted to the momentary needs, regulated by a system where chemoreceptors (peripheral: the carotid and aortic bodies and central: on the ventrolateral surface of the medulla oblongata) are the afferents, and the diaphragm and intercostal muscles the effectors, expanding the lungs. However, respiration is also, partly, under voluntary control and we use compression of the expired air to make sound. This system will even in steady state conditions lead to variability in respiratory rate, partly due to the delays in the feedback loop and partly to changes in 'mood' as a common denominator for a central influence that we don't understand. In both these aspects Respiratory Rate Variability (RRV) and HRV resemble each other, be it on different time scales, 15/min versus 70/min on average. In view of the time delays in the respiratory control loop, the 'Eigen-frequency' of RRV could be around 30–60 s (Karemaker and van den Aardweg 1991). This is also the frequency of occurrence of repetitive sleep apneas or Cheyne–Stokes breathing patterns. A more rigid treatment of the modeling approach to reach these numbers is given in Khoo (2000a) and van den Aardweg and Karemaker (2002). The reason to go into the subject of RRV in an introduction into the ANS is twofold: for one, respiration is tightly coupled to central nervous conditions like 'rest' and 'flight, fight, fright', i.e. parasympathetic or sympathetic predominance. Second, the rhythm of respiration also influences the cardiovascular rhythm and vice versa, as will be discussed next.
2.4.2. Respiratory sinus arrhythmia; cardiorespiratory interaction and the 'whole number ratio'.
During inspiration the alveoli unfold, left atrial pressure drops (Connolly et al 1954) and the pulmonary microcirculation increases its compliance. Consequently, left ventricular output drops and so does blood pressure, causing a baroreflex-mediated increase in HR. As explained in the above discussion on LF/HF this is only a partial explanation of the inspiratory HR-increase of the 'respiratory sinus arrhythmia', i.e. the oscillation in HR accompanying respiration. Another part is the inhibition of vagal motor neurons in the medulla oblongata concurrent with respiration. During expiration, the reverse occurs: left ventricular output increases, so does blood pressure and, via the baroreflex and under influence of respiratory centers, HR decreases. This is a description of how respiration may synchronize heart rate, whereby the two might become phase locked. For this to occur more than just by chance, the alternative pathway must also be active, i.e. inspiration being triggered by the action of the heart, the coupling of the two rhythms leading to what has become known as the 'whole number ratio' between the rates of respiration and that of the heart Something along those lines had very early been observed in experimental animals and was revitalized by Hildebrandt (1953). It became clear that this coupling was very tight during sleep and became variable during the day, depending on the level of attention to the tasks at hand (Engel et al 1969). In more recent years the careful observations by Galletly and Larsen, starting at spontaneously breathing patients under anesthesia (Galletly and Larsen 1999), even if they had atrial fibrillation (Larsen et al 1999). They have elucidated much of this mutual entrainment, leading to the suggestion of an attractive, simple model whereby a gradually increasing inspiratory drive can be helped to cross its action threshold by a burst of afferent activity related to the heart beat. Later, in animal experiments they showed that repeated somatic nerve stimulation could also entrain respiratory onset (Larsen et al 2003), vagotomy did not end it, but baroreceptor denervation did (Tzeng et al 2007). This conclusively proved that the baroreceptor afferent volley was able to trigger the next inspiration.
This concludes the examples of specific organ activity under the action of the ANS. In view of the limitations of this introduction and its author, the reader be referred to specific publications regarding the organs that are not treated here (see table 1). Good overviews of physiology, pharmacology and specific diseases may be found in the textbooks already mentioned (Jänig 2006, Low 2008, Robertson et al 2012, Mathias and Bannister 2013). In particular autonomic nervous control of the GI-tract requires at least a review by itself, if not a book. Of all organs it carries the best resemblance to the 'simple' Hydra anatomy that was discussed above, even the structure of an outer, longitudinal and an inner circular muscle layer seems to have been kept more or less unaltered through evolution.
3. The autonomic nervous system, orchestra or director?
3.1. Global versus local, what do measurements tell?
Our daily life is dominated by autonomic functions, or should I say, bodily functions that occur 'on their own' and drive our actions. We feel hunger or thirst and will try to still those, we urgently must go the toilet, etc. Depending on what we call 'mood' these urges may even be more compelling or strongly depressed. These general observations have led (psycho-)physiologists and medical doctors to try and find numbers to attach to the 'central autonomic condition', by measurement of peripheral functions, since these lend themselves easier to quantification. This presupposes that there is something like a mindset to be characterized by a simple number, in particular one that would indicate the position of the pointer on the dial of sympathetic/parasympathetic balance. This idea explains the big success of the LF/HF number, since it is so easily obtained and claims to have such far-reaching implications. However, it brings in memory the story of the man who was searching for his lost keys in the dark of the night around the only lantern in the street: not because he supposed they would be there, but because that was where the light was shining.
This introduction into the final chapter of this paper may sound a little cynical about the possibilities of drawing conclusions from measurement of autonomic functions. Don't I believe in lie detector tests, didn't I myself measure HRV and LF/HF? No and yes (respectively), but still I think that one should be very cautious in the interpretation of such measurements. Heart rate is, in the first place, a circulatory parameter, and its variability is, indeed, determined by nervous and hormonal inputs impinging on the sinus node, but only after 'translation' by the properties of the pacemaking process. The same will hold for any other measure of autonomic activity, be it pupil size, blood pressure, or sweat production. If the purpose is to 'measure' central autonomic state (if that is possible at all) one should look for central measurements, like fMRI.
Question remains: is there a central autonomic condition which is related to a person's mood and mental drive that we might describe in some way? If so, are the autonomic measures that we extract from peripheral channels in line with this central condition?
3.2. Central versus peripheral autonomic nervous system; a hypothesis
It has been discussed above how the cooperation between ingrowing sympathetic postganglionic neuron and target organ might lead to the specificities of each circuit, as defined by requirements of the effector organ. This might explain how a limited set of preganglionic neurons can successfully command a wide variety of effectors. One may extend this view to the spinal cord where the preganglionic motoneurons are located in the intermediolateral (IML) column of the gray matter, wedged between the dorsal horn where most of the segmental afferent traffic synapses and the ventral horn, the location of the motor neurons for the skeletal muscles. The IML motoneurons receive input from 'higher up' and from segmental somatic and autonomic afferents, making the segmental unit something like a 'subroutine' that can be invoked from higher centers. The clearest examples being those circuits involved in micturition or defecation. For instance, gradual stretch of the urinary bladder is lessened by active smooth muscle relaxation, until the voluntary action of micturition is invoked when a sense of bladder fullness announces itself (in the ideal case of sufficient potty training and continence).
The idea of 'nested subroutines' for autonomic functions may be taken one step further up to the centers above the level of the spinal cord. In its continuation we find the medulla oblongata and the midbrain, together with the pons called the brainstem, the source of 10 out of the 12 cranial nerves (only the olfactory and optic nerves are excluded; strictly speaking the senses of smell and vision are brain functions). Here we also find the autonomic circuits involved in many regulatory functions, like these of the circulation and breathing. At these levels again subroutines can be identified, like that of the beat-by-beat baroreflex blood pressure control (see figure 4) or of the breath-by-breath control of breathing. However, these and the lower, spinal subroutines can also be overridden (or, so you will, be fed with new, situation dependent parameters) if the general conditions require this. Such is commanded from nodes higher-up in the hierarchy: a couple of nuclei at the level of the midbrain itself, but more importantly from the forebrain—the cerebral hemispheres and the structure in between: the diencephalon. Just skipping the brainstem functions, I shall now turn to those forebrain functions, in view of their role as command centers and integrators of somatic and autonomic functions, the nervous and the hormonal systems.
3.3. Integration of autonomic and somatic nervous system; the immune system and the ANS
The best way to consider the integrative mode of action of the various subsystems in the functioning of the individual is by looking during exercise and mental stress. These are also the most-studied conditions in (psycho-)physiology, but many aspects are still not resolved. In a recent review on the muscular hyperemia during exercise Joyner and Casey (2015) picture the microcirculation as a 'sleeping giant' who may take up all the cardiac output if not well-restrained by the sympathetic nervous system. Question remains how this giant is unleashed during exercise: is it by lifting the sympathetic bridle and/or local metabolic factors and/or by active nervous vasodilatation? The arguments in favor of the latter are not strong, but longstanding, in particular supported by what has become known as the 'defense response', i.e. the vasodilatation that may occur during moments of startle or anxiety (Roddie 1977). As mentioned above, in the section on 'neurochemical coding' a multitude of transmitters may be involved in sympathetic activation, not only the classical ones that would favor vasoconstriction.
It is beyond the limitations of this introductory text to even start enumerate all the higher brain nuclei that are supposed to be involved thus far in emotional reactions of the ANS. Be it sufficient to point to the hypothalamic area and parts of the (pre-)frontal cortex (Macefield et al 2013). Another interesting landmark that came up are the 'amygdala', the almond-shaped nuclei that can be found bilaterally hidden in the temporal lobes as extensions of the 'hippocampus', the areas involved in recording of short-term memory. The amygdala are involved in 'coloring' the memories that we make, in particular in coloring them with fear (Izquierdo et al 2016) or the opposite: control of fear in learned behavior (Adhikari et al 2015).
The parallel central command to the exercising muscles and to the ANS will lead to increased sympathetic and decreased parasympathetic outflow, accelerated respiration, increased cardiac output by increased heart rate and mobilization of pooled venous volume. At the same time the hormonal system is switched to the 'active mode' by activating the so-called HPA-axis: from the Hypothalamus to the Pituitary gland to the Adrenal glands a plethora of stress hormones is released, all geared towards increased catabolism, supporting the demands that are made in favor of flight, fight or fright. Furthermore, the immune system is temporarily suppressed.
The communication between the immune system and the ANS has still a lot of hidden pathways of which we have only scratched the surface. A major step was made by the discovery of ACh-receptors on immunocompetent cells, explaining their sensitivity to parasympathetic activation (Nizri et al 2006, Kenney and Ganta 2014). Recently this has led to the development of vagus nerve stimulation as 'electroceutical' for the suppression of rheumatoid arthritis (Koopman et al 2016), which is actually an auto-immune disease, where immune cells are turning against their 'own' colleagues within the same body.
4. Closing remarks
This introduction into autonomic nervous function has been designed to serve the (bio-)physics/-mathematics oriented community. Many of the measurements made in these fields of research are considered just noisy numbers, which are hiding some true, underlying condition. While preparing and writing this paper I became more and more convinced that there are many ways to treat this subject which give the reader just more (new) words but very little new insight. Therefore, I have tried to convey some of my own grasp of the subject, to add some insight to the sheer manipulation of numbers. In my view the measurement noise is part and parcel of the measurement, in some extreme cases the noise is the real signal. In the end, this introduction is a one-man's view, my attempt to help the reader under way and certainly not the flawless last and final word on the functioning of the Autonomic Nervous System.
Footnotes
- 2
A common mistake is to make the plural of 'plexus' as 'plexi'. In Latin the (very confusing) plural of words like plexus and tractus is plexus and tractus. To prevent confusion, I will use an adapted plural, thus plexuses. The same holds for the genitive of these words, which also is plexus and tractus, as in nucleus tractus solitarii, i.e. the nucleus of the solitary tract or NTS, see later.