
Abstract
In the last two decades, it has become evident that the mechanical properties of the microenvironment of biological cells are as important as traditional biochemical cues for the control of cellular behavior and fate. The field of cell and matrix mechanics is quickly growing and so is the development of the experimental approaches used to study active and passive mechanical properties of cells and their surroundings. Within this topical review we will provide a brief overview, on the one hand, over how cellular mechanics can be probed physically, how different geometries allow access to different cellular properties, and, on the other hand, how forces are generated in cells and transmitted to the extracellular environment. We will describe the following experimental techniques: atomic force microscopy, traction force microscopy, magnetic tweezers, optical stretcher and optical tweezers pointing out both their advantages and limitations. Finally, we give an outlook on the future of the physical probing of cells.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Introduction
It is evident that physical forces and mechanical properties play important roles in biological systems, all the way from molecules, single cells to tissues. Prominent processes in which mechanics are essential are for example cell division, cell migration, or muscle contraction. Except for special cases such as muscle, cell mechanics was not systematically and quantitatively studied until the late 1990s. This was mainly because it was difficult to exert and measure the small forces involved in these processes. Typical forces range from pN in the case of molecular motors up to 100s of nN for cell–cell and cell-matrix contacts [1].
Since the first ex vivo cell culture was established roughly a century ago by Harrison [2], cell biology was mainly concerned finding the ideal biochemical culture conditions to mimic the in vivo situation by focusing on temperature, osmolarity, ionic composition, pH, growth factors, and nutrients. Following the work of Harris in 1980 [3], Opas in 1989 [4], and finally Pelham and Wang in 1997 [5] the community realized that substrate elasticity, in particular within the physiological range [6], is an important control parameter for cell behavior and needs to be taken into account [7].
All of the studies mentioned showed that the mechanical microenvironment has a significant impact on cellular homeostasis and that cells actively sense and respond to external mechanics by generating and transmitting forces (e.g. structure formation of contractile acto-myosin stress fibers [8]). To elucidate the complex mechanical interactions with the cells' surroundings, it is important to measure the active and passive mechanical properties of the cell. In the following, we give an overview of the most important of the many techniques that are nowadays used to achieve this goal.
Atomic force microscopy
It was the atomic force microscope (AFM), invented by Binnig et al in 1986 [9], that allowed to exert forces in the pN and nN range on cells in a well-defined fashion. AFM is a surface scanning technique. In contrast to the earlier established scanning tunneling microscope (STM), the AFM can be operated in liquid environments, a crucial requirement for living cells. Samples need to be affixed to a flat surface and a flexible cantilever with a tip as probe at its end is lowered on the sample from above. Laser light is reflected from a cantilever onto a quadrant photo diode and provides a quantitative signal for the deflection d of the cantilever and therefore the applied force F with the help of Hooke's law F = −k·d, with the calibrated spring constant k (figure 1). While the most common use of the AFM is for imaging surfaces with, in the best case, atomic resolution, this makes it possible to also measure mechanical properties of cells, tissues, and hydrogels by indenting them and recording force-distance curves [10]. The applied force versus indentation of the tip of the cantilever (most commonly a pyramid or a sphere) into the sample can be fitted yielding an effective Young's modulus Eeff (see figure 1(B)). Depending on the probe geometry, different mathematical models are used, e.g. for a sphere the modified Hertz model [11], and for a pyramid the cone approximation by Sneddon [12]. If thin layers are probed, such as a flat adherent cell, a correction might be needed as shown by Dimitriades et al [13].

Figure 1. Measurement of micro-elasticity of matrices by AFM. (A) A cantilever with a pyramidal tip (opening angle α) is lowered towards the sample (Δz) and used to indent the sample causing a deflection d monitored by the photodiode, yielding the indentation δ = Δz − d. The exerted force F = −kd is defined by the spring constant k multiplied by the deflection d using Hooke's law. (B) The effective Young's modulus Eeff is determined by analyzing the resulting force-indentation curves with a modified Hertz model. Black curve denotes data points; red dashed line is the best fit resulting from the modified Hertz model. Reprinted from [6], Copyright 2007, with permission from Elsevier.
Download figure:
Standard image High-resolution imageCare has to be taken while fitting data, as the determination of the exact contact point (when the probe touches the sample) can be difficult. The selection of the range of the curve to fit is also a possible source of errors where small deviations can lead to significant changes in the obtained elasticity [14]. A recent paper has focused on combining different strategies to determine the exact contact point [15] to reduce this problem. The other crucial parameter is the spring constant of the cantilever. It is usually calibrated using thermal fluctuations [16], where the power spectral density of the cantilever fluctuations is analyzed. Values obtained can be significantly different from the nominal values that manufacturers report. An overview over several methods to calibrate the cantilever spring constant can be found in this review [17]. Since cells are often very soft, the spring constant of the cantilever should also be low, ideally in the range of 0.01–0.1 nN nm−1. As that implies a relatively low resonance frequency, it is advisable to do the thermal calibration in air to avoid additional uncertainty due to the viscous damping in liquid [18].
While the most common AFM approach to physically probe cells, tissues and hydrogels and determine an effective Young's modulus Eeff is the above described indentation method, it is not straightforward to probe frequency dependent viscoelastic response. Indentation curves are performed at a defined indentation speed and often probe beyond linear response. The frequency dependence of mechanical properties can help tremendously in gaining insight into the structure of complex materials. In bulk rheology, the frequency dependence is commonly probed by applying a sinusoidal stress or strain to the sample and then stepping through frequencies using lock-in detection. A similar approach can be also adapted to be used by AFM as demonstrated by Mahaffy et al [19]. This method was also employed recently to measure differences in the mechanical properties of malignant and benign cell lines where Rother et al showed that cancer cells generally show a larger loss tangent (ratio of loss to storage modulus) than the benign cells [20].
Another way of investigating the frequency dependence of mechanical properties and to separate elastic and viscous contributions is the variation of the speed of force-indentation curves as demonstrated by Nawaz et al [21]. Here, different indentation speeds are used, giving rise to a weak power law dependence of the effective Young's modulus Eeff that significantly depended on the total indentation depth reflecting the heterogeneous structure of cells.
Among other techniques, AFM was also used in a study by Pagliare et al to investigate the mechanics of nuclei of embryonic stem cells at different stages of differentiation [22]. During transition from naïve pluripotency to a committed state, it was found that nuclei exhibit an auxetic behavior where they show a cross-sectional contraction during compression with the cantilever and in addition also an increase of stiffness during compression. Recently, Lherbette et al used AFM microrheology on isolated cell nuclei and could show that there are large spatial inhomogeneities in the structure of the nucleus that might be linked to variation in chromatin organization and could be important for transcriptional regulation via mechano-transduction [23].
In summary, AFM is a powerful technique that besides mapping the 3D topography of samples can also quantitatively measure the mechanical viscoelastic properties. However, it is limited to adherent cells and can only probe from the top of the cell. If simplified models are used to deduce an effective Young's modulus Eeff detailed geometry and typically highly inhomogeneous structure of cells are entirely ignored, and it is often difficult to compare the elastic parameters obtained in this way with those measured from other types of experiments.
Traction force microscopy (TFM)
While AFM can image the topography of cells and measure their mechanical properties by indentation from the apical side, i.e. the side that is not adhering to the substrate, TFM employs the substrate itself to measure forces. TFM records the displacement of beads embedded in an elastic substrate and estimates the transmitted traction force from the basal side of the cell. Dembo and Wang reported for the first time quantitative data for stresses at the cell substrate interface that can amount up to 104 pN μm−2 at a spatial resolution of about 5 µm [24]. Reconstructing the stress field (i.e. the transmitted cellular force to the substrate) from the strain field, sampled via the bead displacements, is a classical inverse problem and as such has its challenges as summarized for example in a review by Bonnet and Constantinescu [25].
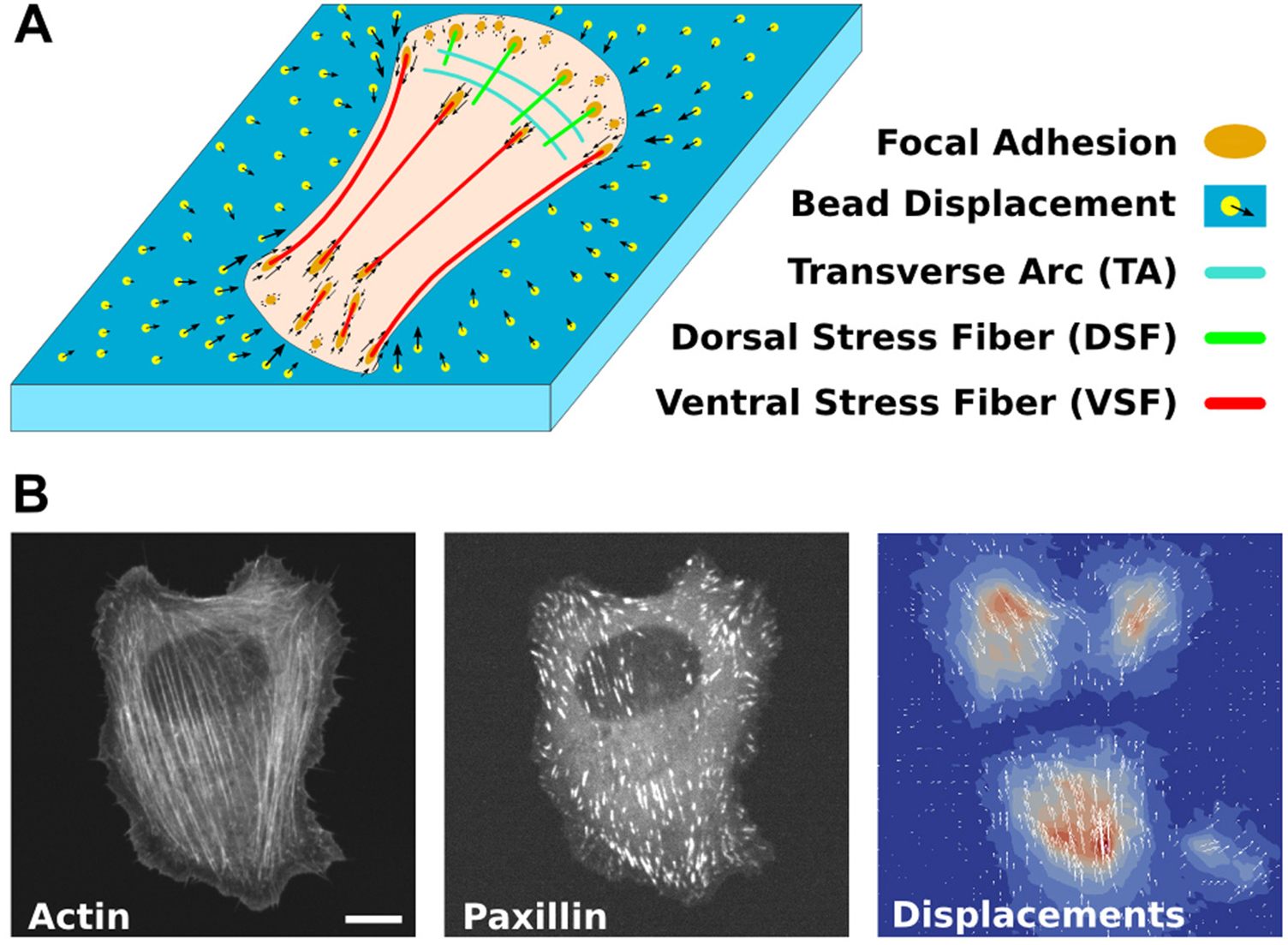
TFM is now well established, and several improvements were developed both on the experimental side and the computational reconstruction of the stress fields [26]. An essential improvement to the solution of the inverse problem of TFM is the use of additional input from microscopy images of the force-transmitting acto-myosin stress fibers of the cytoskeleton and cellular adhesion structures [27] as illustrated in figure 2.

Figure 2. TFM and model-based stress field reconstruction. (A) Sketch of a cell on an elastic substrate transmitting contractile forces from stress fibers via focal adhesions which cause displacements of substrate-embedded beads. (B) Fluorescent micrographs of actin and paxillin in a U2OS-cell and the corresponding bead displacements in the substrate. Scale bar is 10 µm. Reproduced from [27]. CC BY 4.0.
Download figure:
Standard image High-resolution imageThis is a good example of a combination of complementary methods that leads to a better understanding of cellular mechanics than each technique alone could achieve. A recent review summarizes the different computational approaches used, explains the mathematics and also comments on practical issues [28]. Another recent study used TFM to probe the time varying force fields produced by blood platelets [29]. This study was especially challenging since platelets are much smaller than common adherent cells. TFM revealed that platelets exert forces in the range of 30 nN on elastic substrates, significantly higher than previously reported from cell aggregates.
A derivative of TFM is the micro-pillar substrate method. An array of micro-pillars is cast using a silicone elastomer (PDMS), and cells are seeded on top of this 'bed of needles' [30]. Cellular traction forces that are transmitted to the pillars will bend them and by microscopic analysis of the deflection, forces can be obtained. Combining such micro-patterned PDMS arrays with fluorescence microscopy of focal adhesions in living cells that were transfected with a GFP-vinculin fusion protein allows one to precisely determine the local forces at focal adhesions. Surprisingly, a relatively stereotypical value of 5.5 ± 2 nN µm−2 was found. The aforementioned technique was upgraded by Sniadecki et al by adding cobalt nanowires into several posts throughout the whole array of elastomeric posts [31]. This converts the passive pillars to active probes and the 'magnetic' posts can now be deflected by an external magnetic field and probe the cellular reaction to applied forces.
Magnetic tweezers
A geometrically completely different approach to physically probe cells also uses the interaction of magnetic particles with an external magnetic field. Probably the first report on the fundamental principle of magnetic tweezers applied to biological systems was an experiment performed in 1950 by Crick and Hughes that they called the 'magnetic particle method' [32]. Cultured chicken fibroblasts were made to phagocytose ferromagnetic particles. When they applied a magnetic field, Crick and Hughes observed three types of movements of these particles which they described as twisting, dragging and prodding. These experiments clearly demonstrated that the cytosol of cells is not purely viscous but also exhibits elastic properties.
After these pioneering experiments, it took quite some time until the concept of magnetic tweezers was picked up again and further developed. It was only in the 90s that mag-netic tweezers were used to probe single DNA molecules to quantitatively measure force-extension curves and compare with the freely-jointed-chain and the worm-like-chain models [33]. Magnetic tweezers have now become quite popular, in particular for single-molecule force measurements, and are widely used in various implementations (see for example the review by Vlaminck and Dekker [34]).
A crucial issue for these experiments is the precise calibration of the forces acting on the typically paramagnetic beads in the external inhomogeneous magnetic field. Two landmark papers in the late 90s demonstrated precisely controlled forces of up to 10 nN on 4.5 µm beads and probed the mechanical response of adherent cells to the magnetic rocking of beads bound to their membranes [35, 36]. Subsequently, magnetic beads coated with integrin ligands were used to mechanically probe the link between extracellular matrix and the cytoskeleton. By applying forces in the range from pN to 1 nN, Alenghat et al could prove an essential role of the focal adhesion protein vinculin for force transmission [37]. Later, Kollmannsberger and Fabry designed a high-force magnetic tweezers instrument by reducing the distance between the magnet and the beads to a few µm. This made it possible to exert forces up to 100 nN on 5 µm beads, a force that is well-suited to mechanically probe living cells [38]. These forces are in the same range as the forces exerted by cells on their surrounding during matrix adhesion and migration and should make it possible to study the mechanics of such processes in detail. It is crucial in this experiment, however, to bring the magnet extremely close to the probe, which can be difficult for cells in 3D microenvironments or for tissues.
Optical stretcher
The optical stretcher was invented by Jochen Guck and Josef Käs using two opposing divergent laser beams directed on a cell [39]. If both beams have the same intensity there is no net force acting on the cell along the optical axis. Nevertheless, it turns out that, in addition to a lateral optical gradient force keeping the cell on the optical axis, there is a symmetric couple of forces acting on the cell surfaces stretching the cell in the axial direction (see figure 3).

Figure 3. The Optical Stretcher. Two opposing divergent laser beams, delivered by two optical fibers, trap and deform a living cell. The forces acting on the cellular surface can be varied by changing the laser intensity. This makes it possible to estimate the cell's compliance. Reprinted from [39], Copyright 2001, with permission from Elsevier.
Download figure:
Standard image High-resolution imageCells can be trapped in the center of the stretcher and by variation of the laser power, the forces acting on the cellular surfaces leading to the axial stretching can be controlled. An effective cellular compliance J can be extracted from such deformation curves including time dependent information. Such data can be further analyzed in terms of viscous and elastic contributions and can be fitted by either power law rheology models, a standard linear solid model, or a Burger's model [40]. One of the strong advantages of the optical stretcher is the possibility to combine it with microfluidic devices and measure the compliance of cells at a high throughput. This allowed Guck et al to mechanically compare normal, cancerous, and metastatic breast epithelial cells. Distributions of optical deformability were so broad, however, that unambiguous classification of single cells was not possible [41]. Chan et al recently performed measurements on a variety of physiologically adherent and non-adherent cells and also perturbed cell mechanics using biochemical drugs such as blebbistatin, a potent and selective inhibitor of the ATPase of myosin II's, thereby weakening acto-myosin contractility [42]. Interestingly, cells became effectively stiffer with blebbistatin treatment, which is in contrast to measurements by AFM and the dual optical trap [43]. Recently, it was shown that next to the optical deformation of a cell, there is also a significant thermal effect on the sample that allows to induce rapid changes in temperature [44]. Exploiting this thermal effect, the optical stretcher setup therefore allows a new class of experiments called 'thermorheology' that go beyond purely mechanical deformation.
Optical tweezers
Based on focused laser light, optical tweezers can trap and manipulate small refractile objects [45] and are therefore ideally suited for mechanical measurements on proteins and on supramolecular structures up to cells. Seminal experiments using optical tweezers led, for example, to the discovery of the 8 nm steps of the kinesin motor protein [46]. Both displacement resolution and range and resolution of forces ideally match the single molecular necessities. Optical tweezers were also successfully employed to perform mesoscopic measurements of viscoelastic response properties of soft materials [47–49]. These mechanical experiments have been further developed to distinguish local from larger-scale response using one- and two particle rheology of polymer networks [50, 51] and, more recently, to probe the non-equilibrium mechanics of active cytoskeletal networks [52].
Exploiting orthogonally polarized laser beams derived from one laser, one can construct a dual tweezers setup and independently trap and steer two beads (figure 4). Mizuno et al used such a dual optical tweezers to investigate the fluctuations and the mechanical response of suspended cells and could show that the recorded fluctuations were not thermal equilibrium fluctuations, but largely produced by active cellular force generation [53]. A suspended cell is typically mechanically linked via integrins to the two beads that are coated with extracellular matrix ligands (see figure 4(A)). Such a setup can be used not only to record cellular force fluctuations, but also to apply a force to the cell, record the resulting deformation, and by Hooke's law to obtain an effective spring constant.

Figure 4. Dual optical tweezers probing cell mechanics. (a) 3T3 fibroblast suspended between two optically trapped 4 µm fibronectin-coated polystyrene beads. (b) Schematic of the set-up: a cell (grey) exerts forces F1 and F2 onto the two attached beads (red and blue). The corresponding displacements of the beads (u1 and u2) from the trap centers are detected by QPDs and recorded and processed with a computer. In the force-feedback mode, the computer controls an acousto-optic deflector (AOD) that steers one of the traps (blue) to keep the force measured on the other (red) bead constant. (c) Force fluctuations Fi = −k ui of two beads (i = 1, 2) attached to opposite sides of the cell. Reproduced from [43]. CC BY 4.0.
Download figure:
Standard image High-resolution imageSchlosser et al demonstrated that the tensed acto-myosin cortex is affecting the measured cellular effective stiffness. When cells were treated with blebbistatin they got significantly softer, and the transmitted cellular force fluctuations were significantly reduced [43]. This is in contrast to the earlier mentioned results of Chan et al that showed that blebbistatin treatment resulted in a stiffening of cells [42]. An important difference between the two experiments is the attachment of beads coated with matrix proteins that led to stable integrin binding and subsequent focal adhesions for bead diameters larger than 4 µm [54].
When a cell is suspended between two beads, the optical tweezers set the mechanical boundary conditions and, to some extent, take over the role that a substrate plays for adherent cells. In particular, the stiffness of the potential well formed by the optical tweezers can be thought of as roughly equivalent to matrix elasticity. Therefore, a change in laser intensity that affects the trapping potential can simulate a changing mechanical microenvironment. In addition to providing just a passive elastic response, the tweezers can also be used to provide a defined force. It is possible, for example, to keep cells under a constant external load using a feedback mode. Interestingly, when fibroblasts were put under a constant external tension in the range of 5–35 pN they appeared to produce slow oscillatory contractile forces with periods in the range of 1–2 min. This oscillation stopped when cells were treated with blebbistatin that inhibits non-muscle myosin motor proteins, and a creep response to the external load as it is known from visco-elastic materials was observed instead [43].
While the optical stretcher measures the compliance of the whole suspended cell, the dual optical tweezers mechanically couple to the cell via integrin clusters and focal adhesions and transmits forces directly to the acto-myosin cortex. Further information on the cortex and other cytoskeletal structures is needed to fully understand cellular mechanics. This can be achieved in the, compared to adherent cells, simple geometry of close-to-spherical suspended cells by combining the dual optical tweezers with confocal microscopy and potentially appropriate super-resolution microscopy techniques. Figure 5 shows a 3T3 NIH fibroblast in dual confocal tweezers that is transiently transfected with LifeAct-RFP revealing the structure of the acto-myosin cortex but also smaller actin bundles within the cytosol of the cell.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 5. Cell in dual confocal optical tweezers. The left panel shows a phase-contrast image of a 3T3 NIH fibroblast suspended between two beads (diameter 4 µm) trapped in the dual tweezers. In the right panel, the fluorescence image of LifeAct-RFP expressed by the cell shows an equatorial cross section of the acto-myosin cortex and shorter actin bundles in the cytosol, with a region without actin where the nucleus resides.
Download figure:
Standard image High-resolution image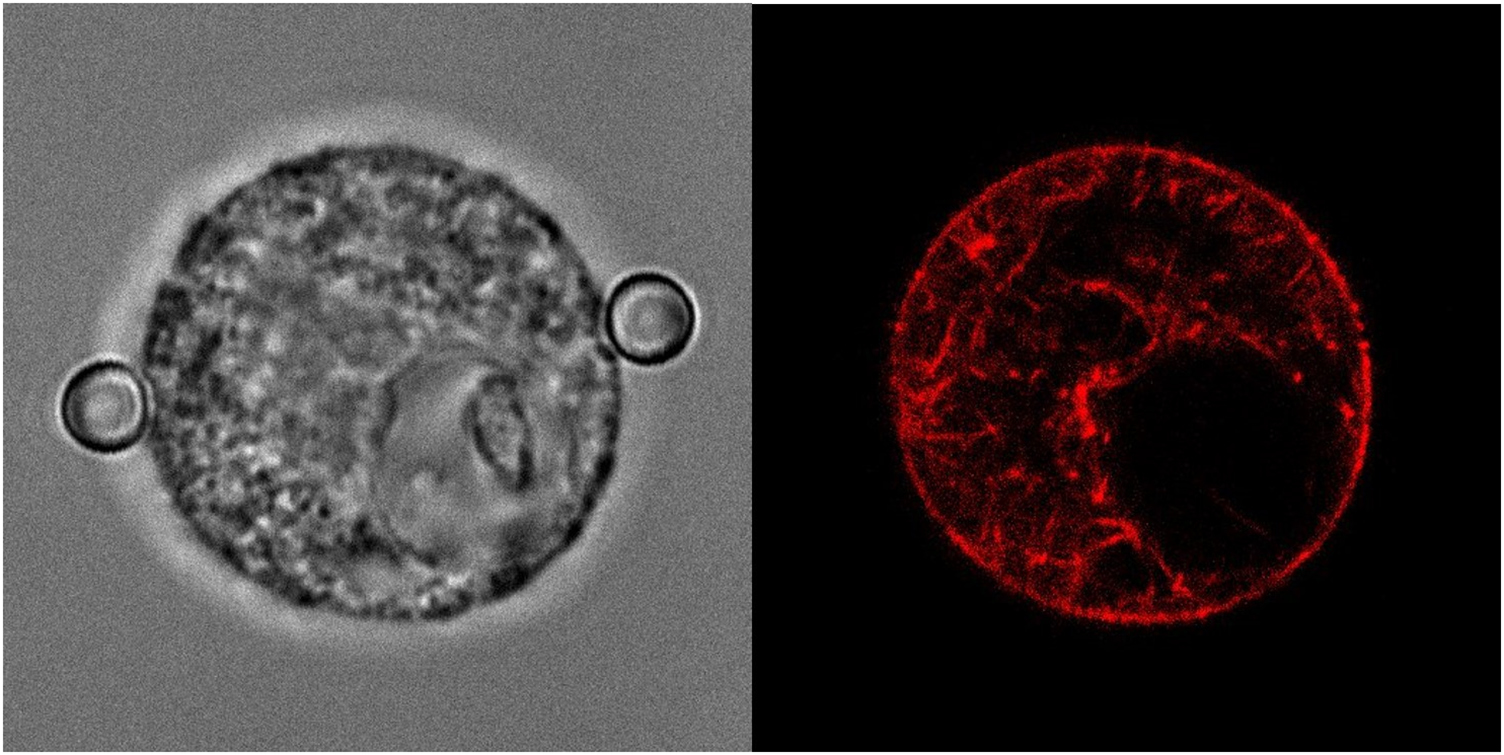{kind=link}
One important limitation for mechanical measurements of cells and eventually cellular aggregates is the limited force range of up to about 100 pN maximal force that optical tweezers can exert. A solution to this problem might be specially designed core–shell particles that showed an increase of maximal trap force by an order of magnitude that might make it possible, together with optimization of optics, to reach forces up to nN [55].
Conclusions and outlook
In this short review, we have highlighted and described a selection of experimental methods that are used to physically probe the active and passive mechanical parameters of cells. Several other methods have been developed (e.g. micropipette aspiration [56], microplate rheology [57], bulk cell rheology [58], real-time deformability cytometry [59], etc). There are two important current applications of physical probing of cells. One is the mechanical phenotyping and distinction of healthy and diseased cells, with a particular focus on cancer diagnostics. Here, many studies with different techniques have already proven their capability of mechanically discriminating between the 'two states', although the distribution of individual properties in any given state is very broad. The other application focuses on a fundamental quantitative understanding and physical modelling of the mechanics of cells in their microenvironment. This area has been rapidly evolving during the last decade but it is fair to say that much still has to be learned.
The development of experimental techniques has gained a lot of momentum in the past decade and will most likely continue to do so. We expect that especially the integration of super-resolution microscopy approaches into existing techniques will promote a new level of understanding. This is important since it will be essential to correlate the active and passive mechanical properties of cells to the corresponding structures that generate and transmit forces.
The experimental results also need adequate physical and mathematical modeling. Although widely used, determining merely an effective Young's modulus Eeff of a cell is an oversimplification and might at best be appropriate to compare two cell types or a knock-out control. Otherwise, the assumption of a homogeneous mechanical continuum cannot account for the heterogeneous complex structures of real biological cells. Here, again, super-resolution techniques will help to elucidate the micro- and nano-scale architectures of cellular structures that are needed to build better models that proceed beyond the simplest approximations. Sophisticated numerical simulations can be used to describe filamentous polymer networks, as models for the cell cytoskeleton, under varying external loads to obtain mechanical properties [60]. There is already compelling evidence that a mean field description of such networks is inappropriate to describe the mechanical response under load since stress focusing on singular elements of the networks, such as stress fibers in cells, occurs even for large networks [61, 62].
Acknowledgments
We acknowledge funding by the German Science Foundation (DFG) via the SFB 937 project A13 and the SFB 755 projects A03. FR gratefully acknowledges support from the Volkswagen Foundation within the Niedersachsen Israel framework (MWK-VWZN2722) and the German Science Foundation (DFG) via the SFB 755 project B08. CFS acknowledges further funding by a European Research Council Advanced Grant PF7 ERC-2013-AdG, Project 340528.