


ABSTRACT
Solar spicules are the fundamental magnetic structures in the chromosphere and may play a key role in channeling the chromosphere and corona. Recently, it was suggested by De Pontieu et al. that there were two types of spicules with very different dynamic properties, which were detected by the space–time plot technique in the Ca ii H line (3968 Å) wavelength from Hinode/Solar Optical Telescope observations. A "Type I" spicule with a 3–7-minute lifetime undergoes a cycle of upward and downward motions; by contrast, a "Type II" spicule fades away within dozens of seconds without a descending phase. We are motivated by the fact that for a spicule with complicated three-dimensional motion the space–time plot, which is made through a slit on a fixed position, could not match the spicule behavior all the time and might lose its real life story. By revisiting the same data sets, we identify and trace 105 and 102 spicules in the quiet Sun (QS) and coronal hole (CH), respectively, and obtain their statistical dynamic properties. First, we have not found a single convincing example of "Type II" spicules. Second, more than 60% of the identified spicules in each region show a complete cycle, i.e., the majority are "Type I" spicules. Third, the lifetimes of the spicules in the QS and CH are 148 s and 112 s, respectively, but there is no fundamental lifetime difference between the spicules in the QS and CH reported earlier. Therefore, the suggestion of coronal heating by "Type II" spicules should be taken with caution.
Export citation and abstract BibTeX RIS
1. INTRODUCTION
The spicule was initially discovered by Secchi (1877) of the Vatican Observatory in Rome, Italy, and later was named as a spicule by Roberts (1945). It has a jet-like luminous structure for its long and slim profile (Beckers 1972; Lorrain & Koutchmy 1996). Generally, the spicule could be seen through chromospheric lines, such as Hα, Hβ, D3, Ca ii H, and K (Michard 1954). In recent decades, larger-size spicules with similar structures were observed in  , ultraviolet (UV), extreme-ultraviolet (EUV), and soft X-ray wavelengths, called UV, EUV, and soft X-ray macrospicules, respectively (Bohlin et al. 1975; Dere et al. 1989; Wilhelm 2000; Xia et al. 2005).
, ultraviolet (UV), extreme-ultraviolet (EUV), and soft X-ray wavelengths, called UV, EUV, and soft X-ray macrospicules, respectively (Bohlin et al. 1975; Dere et al. 1989; Wilhelm 2000; Xia et al. 2005).
A huge number of spicules, like messy hair, and the interspicule are composed of the chromosphere. The mass flux taken by the spicules to the corona is exceeding that of solar wind by two orders of magnitude (Thomas & Athay 1961). Therefore, besides working as the indication of an inhomogeneous chromosphere, the spicule is thought to be a very likely candidate for transporting material and kinetic energy into the corona as well as heating it (Woltjer 1954; Rush & Roberts 1954; Li & Ding 2009). Because of its mysterious formation mechanism, and the possibility that it may heat the corona, it has always attracted the strong attention and interest of researchers. Suematsu et al. (1982) suggested that the spicules are formed as a result of slow-mode shocks propagating along vertical magnetic flux tubes in the photosphere and low chromosphere by using one-dimensional hydrodynamic simulation. Shibata & Suematsu (1982) explained why spicules are taller in the coronal hole (CH; Beckers 1972) by extending the slow-shock model. Later, the Alfvén wave model was successfully proposed to explain spicules and their role in heating the corona (Hollweg et al. 1982; Sterling & Hollweg 1988; Hollweg 1992; Kudoh & Shibata 1999). However, it should be noted that even in the Alfvén wave model, slow shocks are generated due to nonlinear mode coupling with Alfvén waves and play an essential role in the acceleration of the spicules (Saito et al. 2001). De Pontieu et al. (2007b) discovered ubiquitous Alfvén waves on spicules using Hinode/Solar Optical Telescope (SOT). A ubiquitous horizontal field discovered in the photosphere (Lites et al. 2008; Ishikawa et al. 2008; Jin et al. 2009; Zhang et al. 2009) might a trigger reconnection in the photosphere and the low chromosphere, which may excite Alfvén waves (Takeuchi & Shibata 2001; Isobe et al. 2008). Based on the model proposed by Suzuki & Inutsuka (2005, 2006), by considering the observed photospheric granular buffeting as the source of Alfvén waves, Matsumoto & Shibata (2010) successfully reproduced spicules, corona, and solar wind.
Previous observations of individual spicules were difficult because of the low observation resolution (Sterling 2000). The situation has been much improved since the construction of Swedish 1 m Solar Telescope (Scharmer et al. 2003) in 2003 and the launch of Hinode satellite (Tsuneta et al. 2008; Suematsu et al. 2008a; Ichimoto et al. 2008) in 2006. Hence, it is meaningful to remeasure the dynamic properties of the spicules with these free-seeing data sets and to further study the coronal heating. According to the new observations, De Pontieu et al. (2007a) claimed that the spicules should be divided into two types: "Type I" spicules with a 3–7-minute lifetime that are driven by shock waves formed as a result of p-mode leakage; and "Type II" spicules, which are a result of magnetic reconnection, bear a much shorter lifetime of about 10–150 s and a faster speed between 50 and 150 km s−1. The "Type II" spicules dominate the structure of the solar chromosphere in the CH. Without a downward moving phase, "Type II" spicules promptly fade away into the corona (De Pontieu et al. 2007a; Sterling et al. 2010), so it seemed natural to accept this mechanism to explain the coronal heating.
Yet, from the filtergrams, the movements of the spicules could be observed in both horizontal and vertical directions. If spectral observations were available, we could achieve the Doppler shift in the line of sight (Nikol'Skii & Sazanov 1967; Pasachoff et al. 1968; Weart 1970; Suematsu et al. 1995). Spicule movement usually appears in a more complicated, three-dimensional (3D) way, but we do not know what its real movement is. The movements observed in filtergrams are thought to be related to the waves, such as kink and Alfvén waves (De Pontieu et al. 2007b), and oscillations (Kulidzanishvili 1983). We notice that the space–time plot, which could reflect the dynamic information of the spicule to a certain extent (Banerjee et al. 2000; Christopoulou et al. 2001; Tavabi et al. 2011), are always used in measuring the lifetime and height of spicules. For the real 3D motion of a spicule as mentioned above, especially the extensive lateral movement, a spicule cannot always keep its motion in a fixed direction. Therefore, some doubts arise on the reliability of the lifetime and height measured by the method.
We are motivated, therefore, to remeasure the dynamical properties for a better understanding of the spicule model. In this work, the data sets in De Pontieu et al. (2007a) are revisited. We first re-examine types of morphology that were regarded as being spicule by De Pontieu et al. (2007a). Are they real spicules? If not, what is the distinction between them and those spicules directly observed through filtergrams? By drawing a comparison on the height and lifetime, it is discovered that the "spicules" identified in the space–time plots usually have shorter lifetimes and lower heights than those in the filtergrams. Then, to show the statistical properties of the spicules, we trace 105 and 102 spicules in the quiet Sun (QS) and CH, respectively, and survey their distributions of lifetime, height, velocity, and acceleration.
Section 2 will introduce the observations and method adopted in this paper. The results will be listed in Section 3. The discussion and conclusion are in the last section.
2. OBSERVATIONS AND ANALYSIS
2.1. Data
We use the same data sets in QS and in CH adopted by De Pontieu et al. (2007a). The observations were carried out by the Hinode/SOT Broadband Filter Imager in Ca ii H filter whose bandwidth is broad enough to simultaneously observe both the photosphere and chromosphere.
The selected QS, near an α-type active region, NOAA 10923, is located in the western limb of the Sun with the center coordinates of 960'', −90'' and the field of view (FOV) of 56'' × 56''. From 00:00:04 UT to 02:19:59 UT on 2006 November 22, there are a total of 1050 frames observed with a pixel size of 0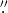 05 and time cadence of about 8 s. The center coordinates of the CH are of 0'', −968'' with the same size of FOV as the QS. A total of 758 frames are available in observations from 11:29:32 UT to 12:30:00 UT on 2007 March 19. The spatial resolution remained the same but the temporal resolution was improved to 5 s or so. The IDL routine in the libraries of Solar Software, fg
05 and time cadence of about 8 s. The center coordinates of the CH are of 0'', −968'' with the same size of FOV as the QS. A total of 758 frames are available in observations from 11:29:32 UT to 12:30:00 UT on 2007 March 19. The spatial resolution remained the same but the temporal resolution was improved to 5 s or so. The IDL routine in the libraries of Solar Software, fg prep.pro, is applied to the image reduction to correct for dark currents and other errors of the camera. Then, we remove the cumulative offsets and have the data set co-aligned. Additionally, we use the radial density filter (Okamoto et al. 2007) to enhance the visibility of the spicules.
prep.pro, is applied to the image reduction to correct for dark currents and other errors of the camera. Then, we remove the cumulative offsets and have the data set co-aligned. Additionally, we use the radial density filter (Okamoto et al. 2007) to enhance the visibility of the spicules.
2.2. Comparison of the Spicules Observed Using the Two Methods
In the space–time diagrams, a total of five typical morphology appearances of "spicules" have been identified with large discrepancies of their lifetimes and movement modes as classified by De Pontieu et al. (2007a). The "Type I" spicules are indicated by "A" and "B" in Figure 1 and the "Type II" spicules by "C" and "D" in Figure 1 and by "E" in Figure 2. The tops denoted by triangles in Figures 1 and 2 are determined by the space–time plot at seven points of time for each case.

Figure 1. Comparison of two methods for tracking spicules. In the panels of the first column are four typical morphology appearances of "spicules" determined by the space–time plots. The second column shows the height–time plots. The last seven columns are Ca ii H filtergrams observed by Hinode/SOT. The triangle denotes the height acquired by the space–time plot; as a comparison, the plus sign is the height obtained by frame tracking directly.
Download figure:
Standard image High-resolution image
Figure 2. Same as Figure 1 but for the fifth typical morphology appearance of the spicule. The first panel in the top left shows the spicule identified by the space–time plot; the second panel shows the height–time plots; the third and fourth panels in the bottom right show the Ca ii H filtergrams observed by Hinode/SOT. The triangles and the pluse signs are the same as in Figure 1.
Download figure:
Standard image High-resolution imageYet, it is uncertain whether the true height is obtained or not. To ensure that it has, we try to find the tops of the "spicules" in the filtergrams. In Figure 1, starting with the third panel, its seven panels in the row are the filtergrams corresponding to the seven points of time. The plus sign indicates that the true height of the spicule is directly determined by the filtergrams. The scatter plots in Figures 1 and 2 are used to compare the positions of the tops acquired by two methods, respectively. For example, in the panel marked with "A" in Figure 1, the plus signs basically match those triangles. It means that, in this case, the lifetime and height determined by the space–time plot is basically true. However, in the case of "B," the triangles are always slightly lower than the plus signs, although only several pixels apart between the slit for the space–time plot in the example "B" and the one in the example "A." The errors of the height measured in the space–time plot would produce errors in velocity and acceleration as well. In the sample "C," we find that there is also great discrepancy in height measurement between the two methods. In the space–time plot, the "spicule" seems to fall off suddenly; however, according to the filtergrams, the spicule falls more slowly and does not finish its entire life experience, as shown in the corresponding scatter plots. The example "D" is a more "typical" "Type II" spicule in the space–time plot, but, in the filtergram method, the undetected descent phase by the first method did exist. As far as the last example "E" (shown in Figure 2) having a strange shape, the reason why it lacks the ascent phase is that the first half of the process was not yet recorded in the space–time plot. The entire lifetime should include the panels enclosed by the dotted frame, indicating the ascent phase and the dashed frame, the descent phase, but the space–time plot failed to detect the first part. Morphologically, in the space–time plots, as has been pointed out by De Pontieu et al. (2007a), two types of spicules are identified. But, the approximately vertical stripes ("Type II" spicule) may be caused by employing the slit, which simply cuts part of the brightness distribution of a spicule, so that only a strip of the distribution is left to be shown in the plots. For this reason, we hope to achieve a statistical result of the spicule dynamic properties to determine whether there are two types of spicules and their distributions in QS and CH.
2.3. Measurement of the Kinetic Parameters of the Spicule
In the data set of the QS, we identify 36, 33, and 36 spicules in three frames, i.e., Frame No. 100, 300, and 500, respectively, for a total of 1050 filtergrams observed by the Ca ii H filter. The upper panel of Figure 3 shows the No. 300 frame shot in the QS at 00:40:05 UT on 2006 November 22. The tops of the 33 identified spicules (numbered 37–69) in this frame are marked by small squares in five colors: red, green, blue, purple, and yellow. Meanwhile, their serial numbers are also written in the same color over the indicated spicules. The white dashed squares illustrate the FOVs of those filtergrams in Figures 4 and 5.

Figure 3. Ca ii H images acquired by Hinode/SOT for the quiet Sun at 00:40:05 UT on 2006 November 22 (upper panel) and the coronal hole at 11:41:31 UT on 2007 March 19 (lower panel). Those colorful minisquares mark the tops of identified spicules numbered with the concolorous Arabic numerals. The yellow lines denote the positions of slits to make space–time plots; white dashed frames are the field of view of filtergrams in Figures 4 and 5, respectively.
Download figure:
Standard image High-resolution image
Figure 4. Time series of the dynamic processing of No. 55 SQ observed by Hinode/SOT Broadband Filter Imager in the Ca ii H on 2006 November 22. In each subimage, the red triangle marks the top of the spicule. The green solid curve is the brightness variation along the green dotted line. The red dashed line denotes the brightness value of 20. Similarly, the red solid curve is the brightness variation along the red dotted line. Two upper and ten lower cyan solid curves are also brightness variations along corresponding lines that are parallel to the red dotted line, but they are upper and lower than the red dotted line, respectively. The panel at the bottom right is the height–time plot of this spicule.
Download figure:
Standard image High-resolution image
Figure 5. Time series of the dynamic process of No. 17 SC observed by Hinode/SOT Broadband Filter Imager in the Ca ii H on 2007 March 19. The lines have the same meaning as those described in Figure 4. The bottom right panel is also the height–time plot of the spicule.
Download figure:
Standard image High-resolution imageFigure 4 represents the time series of the dynamic process of the No. 55 spicule in the QS (hereafter referred to as SQ 55). The spicule, as shown from the third panel to the third last panel, is detected from 00:39:57 UT to 00:43:17 UT. To find the top of the spicule at each time, based on a naked-eye view, we used a set of brightness curves as an auxiliary measure to acquire a more precise position for each top. These brightness curves reflect the brightness variation along the 13 horizontal lines with a 2-pixel distance separating the two adjacent lines. When the lines covered the spicule SQ 55, the brightness curves are plotted exactly above the spicule. Among them, the red brightness curve displays the brightness variation of the red dotted horizontal line. A spine shape gradually forms, becoming quite evident at about 00:41:09 UT, and then slowly weakens. At 00:43:17 UT, it was almost undetectable because it dropped to a lower position and then mixed with other spicules. The upper end of the spine, which is almost as weak as its ambience, should be the position of the top of the spicule at each moment. A series of red triangles corresponding to the time are used to indicate the spicule top positions. A green dotted vertical line and its concolorous brightness curve are plotted as a reference. The brightness has an apparent variation around the top. A long red dashed vertical line marks the brightness value of 20 in the vertical direction. In the bottom right panel in Figure 4 is the height–time plot, and the spicule height at each moment determined by the filtergrams is shown with the diamond. The apparent lifetime and height of the spicule SQ 55 are 200 s and 4819 km, respectively. The top trajectory marked by the diamonds shows a typical parabolic profile covering a complete cycle of ascent and descent. Before 00:39:57 UT and after 00:43:17 UT, due to the overlapping by a very large number of spicules, the SQ 55 could not be exactly detected. Evidently, the real lifetime should be longer than the apparent one, although we could not determine the real path of the spicule in which it was overlapped by other spicules. In total, as shown in Table 1, there are 71 of 105 (67.6%) spicules in the QS moving upward first and then falling back. As for the other 34 spicules, as shown in Figure 6, it seems that there are three different types of spicules. Only the first "type" has the ascent phase, indicated by SQ 82; the second as SQ 33 descends from higher to lower; and the last one, denoted by SQ 44, shows no obvious ascending or descending behaviors. Thus, the profiles of these three types are likely the result of trajectory mixture with surrounding spicules. We still could not trace the entire life for each individual spicule.

Figure 6. Time series of the dynamic process of the other three types of spicule and the related height–time plots. No. 82 is only observed at the ascent stage, No. 33 at the descent, but No. 44 spicule shows no obvious change in its height.
Download figure:
Standard image High-resolution imageTable 1. Statistical Dynamic Properties of Four Types of Spicules
| QS | CH | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Properties | No. | H | LF | Vy | a | No. | H | LF | Vy | a |
| (km) | (s) | (km s−1) | (km s−2) | (km) | (s) | (km s−1) | (km s−2) | |||
| Up- and downward | 71 | 5174 | 176 | 16.9 | −0.13 | 64 | 9572 | 121 | 48.0 | −1.37 |
| Only upward | 9 | 4792 | 74 | 11.2 | −0.10 | 24 | 10,391 | 100 | 26.7 | −0.22 |
| Only downward | 11 | 4729 | 82 | −9.6 | 0.05 | 6 | 8176 | 86 | −6.7 | −0.39 |
| Uncertainty | 14 | 4570 | 103 | ... | ... | 8 | 8418 | 95 | ... | ... |
| Sum(mean) | 105 | 5014 | 148 | 15.5 | −0.14 | 102 | 9592 | 112 | 40.5 | −1.04 |
Note. No. H, LF, Vy, and a refer to the number, the mean height, the mean lifetime, the mean velocity in the vertical direction, and the acceleration in the vertical direction, respectively.
Download table as: ASCIITypeset image
Similar to the data set of the CH, from Nos. 150, 300, 450, and 600 frames, 20, 24, 34, and 24 spicules have been identified and traced, respectively. The lower panel of Figure 3 shows the No. 150 frame with 20 identified spicules that have been indicated with the same symbols as above. The added white dashed frame is similar to the illustration of the FOV used in the filtergrams of Figure 5. Figure 5 shows the entire tracing process of the No. 17 spicule in the CH (hereafter referred to as SC 17) from 11:40:43 UT to 11:44:38 UT, i.e., its apparent lifetime being 235 s. In contrast with the spicules in the QS, the peaks of the spicules in the CH are too dim to be accurately detected. In the left lower panel of Figure 5, the tops at each time, determined by frame tracing, are shown in the height–time diagram. Apparently, SC 17 also experienced a complete cycle, e.g., up and then down. As is listed in the right column of Table 1, a total of 102 spicules in the CH, 64 (62.7%), 24 (23.5%), and 6 (5.9%), have acquired the upward and downward, only upward, and only downward phases, respectively; the remaining eight (7.9%) spicules have no apparent upward or downward phase.
2.4. The Dynamic Properties of the Spicules in the QS and CH Regions
By tracing 105 and 102 spicules in the QS and CH, we obtain the histograms of height, lifetime, and vertical velocity shown in Figure 7. In panels A and B, the mean height in the QS is 5014 km, ranging from 1027 to 8690 km, which is much lower than 9592 km, compared with 4819 to 17,142 km in the CH. The average mean apparent lifetime in the QS is 148 s, while in the CH it is 112 s. According to the height–time plot of each spicule in the QS and in CH, we find that there are 71 out of the 105 (about 67.6%) spicules in the QS and 64 out of the 102 (about 62.7%) in the CH that have a relatively complete cycle of ascent and descent movement. The reason those spicules do not have a cycle basically lies in the disorder of overlapping a huge number of spicules. Hence, for some spicules, we observed the entire process of rising from the back of their adjacent spicules and then falling down, and, for others, we only observed ascending, descending, or an intermediate stage between. Therefore, the measured lifetime should be shorter than their real lifetime. Yet, it is not possible to tell what will occur when a spicule submerges into its background. Additionally, the velocity is basically proportional to the deceleration, both in the QS and in CH (see panels (G) and (H) in Figure 7).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 7. Histograms for QS (2006 November 22) and CH (2007 March 19). (A) and (B) are the maximum height, (C) and (D) are the lifetime, (E) and (F) are the maximum vertical velocity, and (G) and (H) are the scatter plot of the spicule's maximum velocity vs. its maximum deceleration in the vertical direction.
Download figure:
Standard image High-resolution image{kind=link}
3. DISCUSSION AND CONCLUSION
For the lateral motion of a spicule either in the QS or in CH, no fixed slit could be employed during its entire lifetime. Therefore, the spicule profile formed from the space–time plot could not exactly represent the real trajectory of its movement. Comparing the kinematic parameters of the spicules obtained in both methods, the height and the lifetime acquired with the first method are always much less than those with the method we have adopted. That is why we do not use the space–time plot but directly employ its filtergrams to trace the trajectory of each spicule. More than 60% of the spicules move in a complete cycle of ascent and descent in the QS and CH, and the rest do not show a cycle mixed in the background. In brief, with the same data sets in the QS and in CH already used by De Pontieu et al. (2007a), no convincing Type II spicule has been captured. Therefore, Type I, but not Type II, spicules dominate in the QS and CH as shown in Table 1. This result is consistent with the study of spicules in the disk over the QS (Suematsu 1998), over a plage (Anan et al. 2010), and at the limb (Pasachoff et al. 2009). It suggests that there is not a sufficient number of Type II spicules to heat the corona because they fade away in both the QS and CH.
Surely there are apparent discrepancies in the dynamic properties of spicules in the QS and in CH. The spicules in the CH seem to be more energetic than those in the QS (Shibata & Suematsu 1982); for instance, they hit at a much higher position with higher speed and their lifetime is shorter. However, in both the QS and CH, their velocities are approximately proportional to their accelerations, respectively. There is a kind of faster spicule, but this does not mean that its physical mechanism is essentially different from that of other spicules.
With the help of the unprecedentedly high spatial resolution of the Hinode/SOT Ca ii H filter, the identified diameter of a spicule could be less than 200 km, which is much thinner than the values observed in the past. However, it is still not easy to acquire a complete image of a limb spicule. Because of the faintness at the top of the spicule, there is some uncertainty in determining the top of a spicule (Rush & Roberts 1954). At the lower position, as Lynch et al. (1973) pointed out, the spicules are very crowded; only when the visible spicules rise to some height and further separate from each other could we measure their kinetic parameters (Woltjer 1954). This means that the measurement of the dynamic properties of spicules is far from perfect. Thus, the life story of spicules remains somewhat vague. New instruments and techniques are reasonably expected.
Last, but not least, when we watch the animation of the Ca ii H filtergrams, spicules may form a group, moving in a similar way, such as dancing a waltz or as a bamboo raft dispersing into individuals, etc. The group motion may be a more complicated behavior of a double-thread structure of a spicule with the following evolution, i.e., expansion thread separation, lateral motion and spinning as a entire body (Suematsu et al. 2008b; Sterling et al. 2010), which was speculated to be magnetic reconnection. Thus, besides the energy taken by the spicules themselves, these intensive activities will carry a huge amount of energy from the chromosphere into the corona. It may be the essentials of a macrospicule and the practical way of releasing energy to the corona. Our next work will pay more attention to measuring the kinematic parameters in 3D combined with ground-based Doppler observations in the line of sight (Shoji et al. 2010).
Hinode is a Japanese mission developed and launched by ISAS/JAXA, collaborating with NAOJ as a domestic partner and NASA and STFC (UK) as international partners. Scientific operation of the Hinode mission is conducted by the Hinode science team organized at ISAS/JAXA. This team mainly consists of scientists from institutes in the partner countries. Support for the postlaunch operation is provided by JAXA and NAOJ (Japan), STFC (UK), NASA, ESA, and NSC (Norway). We are grateful to the anonymous referee for the insightful comments and suggestions which helped us considerably improve the manuscript. The discussion with Dr. H. Isobe, Dr. P. F. Chen, H. Watanabe, T. Kawate, and T. Anan is very appreciated. This work is supported by the Young Researcher Grant of National Astronomical Observatories, Chinese Academy of Sciences (118900KX3), the National Natural Science Foundations of China (40890161, 40974112, 11025315, 11073050, and 11003026), the CAS Project (KJCX2-EW-T07), and the National Key Basic Research Science Foundation Program of China (2011CB811403) and Y07024A900.