


Abstract
The response of plant diversity to increased snowfall, i.e., precipitation that falls in a solid state rather than a liquid state, is unclear. We investigated the potential effects of 12 year snowfall augmentation on species richness using coordinated distributed experiments, including ten sites across a rainfall gradient of 211–354 mm and spanning 440 km in length in the temperate steppe. Snowfall augmentation decreased species richness rather than enhancing it. Abiotic factor driven by soil pH was the dominant determinant affecting the variation in species richness under changing precipitation regimes, overriding biotic factor. The strongest reduction in species richness induced by snowfall augmentation occurred in the low-rainfall sites. Our study provides insights into the relationship between precipitation and biodiversity in arid and semiarid regions.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Introduction
Global climate change not only alters rainfall regime during growing season, but also changes snowfall pattern within non-growing season (Knowles et al 2006). Increased rainfall enhances plant diversity in arid and semiarid regions by promoting variability in soil moisture (Yang et al 2011), and/or by decreasing the effects of species interactions on species diversity (Gaston 2000). However, increased snowfall may have different impacts on plant diversity from increased rainfall due to their different impacts on surface albedo, soil temperature and evapotranspiration (Screen and Simmonds 2012, Berghuijs et al 2014).
Increased snowfall could impact plant diversity by altering the abiotic factors (Laliberte et al 2013), such as shortening the growing season (Jonas et al 2008), elevating the winter soil temperature (Brown and DeGaetano 2011), and increasing the spring soil moisture (Litaor et al 2008). First, the shortening of the growing season length was shown to decrease species richness through reducing energy availability for plant (Carlson et al 2015). Second, warmer soil under the insulating layer of deep snow exhibits increased organic matter decomposition during winter, which could enhance the availability of nitrogen to plants in the subsequent growing season (Buckeridge and Grogan 2008). However, soil inorganic nitrogen (SIN) enrichment often causes declines in species richness (Phoenix et al 2012). Third, increased soil moisture may partially alleviate the water stress that plants experience before the arrival of rainfall, which enhances species richness (Yang et al 2011). Therefore, elucidating the abiotic effects of increased snowfall on species richness will facilitate the mechanistic understanding of plant diversity in response to precipitation changes.
Increased snowfall can also change species interactions, which serve as biotic factors influencing species richness. First, a shortening of the growing season under an increased snowfall regime leads to a longer period of overlap during which species compete for limited resources, which results in negative neighbor effects (Wipf et al 2006). Second, increased snowfall alleviates soil water and nitrogen deficiencies, which shifts below-ground competition for soil resources to above-ground competition for light (Chesson et al 2004). In such cases, high-stature species possess a competitive advantage over low-stature species through shading effects (Yang et al 2011), which also results in negative neighbor effects (Hautier et al 2009). Therefore, distinguishing the abiotic versus biotic factors effects facilitates a mechanistic explanation of changes in plant diversity under changing precipitation regimes.
Plant diversity in arid and semiarid regions is tied heavily to snowfall during the non-growing season and rainfall during the growing season (Prevéy and Seastedt 2014). However, it is not known whether the magnitude of the species richness response to snowfall at the site-scale will be altered by rainfall at the regional scale. Increasing evidence has shown that ecological sensitivity to resource availability varies markedly across a rainfall gradient (Huxman et al 2004, Huston 2012). For example, rain-use efficiency, the ratio of above-ground net primary production (ANPP) to rainfall, decreased significantly in wetter sites (Bai et al 2008). In contrast, ecosystem stability, the opposite of inter-annual variation of ANPP, increased progressively in wetter sites (Bai et al 2007). Although increased precipitation generally increases ANPP in water limited environments, it remains unclear how increases in precipitation in water limited environments, especially as snowfall, affect plant diversity.
Located in the eastern part of the Eurasian grassland biome, the semiarid steppe in northern China has an area of 313 million hm2 and a plant species richness of 2300 (Kang et al 2007). Observational studies and model simulations are predicting an increase in mean snowfall and extreme 'snow disaster' snowfall in this area (Peng et al 2010, O'Gorman 2014). To understand the general effects of increased snowfall on plant diversity, we conducted coordinated distributed experiments (Fraser et al 2013), including ten sites across a rainfall gradient of 211–354 mm and spanning 440 km in length in Inner Mongolia. The snow fences in our study were installed at each site in 2003. The specific questions addressed were as follows: (1) How does increased snowfall affect plant diversity in arid and semiarid regions? (2) Do drier and wetter sites show a similar snowfall sensitivity of plant diversity?
Materials and methods
Study area and experimental design
This study was conducted in the Inner Mongolia grasslands in the central Eurasian steppe (figure 1). The study area is located at 43.46 °N–45.83 °N latitude, 115.78 °E–119.72 °E longitude, and the elevation is between 908 and 1257 m (table S1 is available online at stacks.iop.org/ERL/13/094002/mmedia). The mean annual precipitation (MAP) ranges from 228 to 379 mm, approximately 8% of which falls as snow. The mean annual temperature ranges from 1.32 °C to 3.10 °C, with the lowest mean monthly temperature in January and the highest in July.

Figure 1. Study area and locations of the ten increased snowfall experimental sites. Coordinated distributed experiments were conducted across a mean annual rainfall gradient from 211 to 354 mm in the temperate steppe in Inner Mongolia, China. Satellite image of Asian snow cover is from National Aeronautics and Space Administration. Individual site names are abbreviated: AER, Aershan; DON, Dongwu; Bayanbaolige, BAY; Wulagai, WUL; XIL, Xilinhot; BAI, Baiyinxile; DAL, Dalinuoer; HAL, Halagaitu; GAN, Ganqiaobao; HUO, Huolinguole.
Download figure:
Standard image High-resolution imageCoordinated distributed experiments were established across a rainfall gradient, which spans approximately 440 km in length. The experiments consisted of ten sites covering two major vegetation types: typical steppe and meadow steppe. The typical steppe, located in the western part of the rainfall gradient, is dominated by Leymus chinensis (Trin.) Tzvel., Carex enervis C A Mey., Cleistogenes squarrosa (Trin.) Keng and Stipa grandis P Smirn. and has lower species richness and productivity. The meadow steppe, located in the eastern part of the rainfall gradient, is dominated by Carex enervis C A Mey., Leymus chinensis (Trin.) Tzvel., Cleistogenes squarrosa (Trin.) Keng and Agropyron cristatum (L.) Garrtn. and has higher species richness and productivity. Soil types include typical chestnut and dark chestnut, corresponding to typical steppe and meadow steppe, respectively.
Along the rainfall gradient, a total of ten snowfall augmentation experiments were conducted. Snow fences were 60 m long and 1.8 m high and installed at each site as part of road snow control efforts by the Inner Mongolia Department of Transportation in 2003. The snow fences were generally perpendicular to the prevailing wind direction in winter, which resulted in equilibrium snow drifts accumulated on both sides of the fence. The method of snowfall augmentation has been used to study the effects of increased snowfall on ecosystem functions in recent studies (Chimner et al 2010, Loik et al 2013). The snow fences range from 40 to 60 m away from the ditch of the highway. The study plots were located within the fenced buffer zone along the road, which has not been grazed by large herbivores for more than 12 years.
The snow fence created zones of differing snow depth (figure S1), including mildly, moderately, and heavily snowfall augmentation, at approximately 10 m, 7 m and 3 m upwind of the fences, respectively. The heavily snowfall augmentation were within the range of a large 'snow disaster' snowstorm (Peng et al 2010). The control plots were established at approximately 30 m away from upwind of the fences where snow depth was unaffected by the fence. We established 3 vegetation plots for each level of snowfall augmentation treatments as replicates, resulting in a total of 12 plots for each site. Snow depth in each plot was measured using poles pushed through the snow to the soil surface during 1–5 February 2015, corresponding to the annual peak snowpack depth. Snow water equivalent in each plot was a product of snow depth multiplied by snow bulk density (0.13 g cm−3 for northern China) (Mo et al 2016). We identified that 0.13 g cm−3 is an assumption, which has uncertainty, although the assumption method has been accepted in recent studies (Hu et al 2013).
Field sampling and measurements
Field sampling was carried out during 15–25 August 2015, corresponding to the annual peak in standing biomass (Bai et al 2008). Species richness was estimated using the number of species found in each 1 × 1 m quadrat. For each species, the number of individuals was measured before clipping. For each quadrat, live vascular plants were clipped at ground level and sorted into species. The above-ground biomass (AGB) in each 1 × 1 m quadrat was determined as the weight of the above-ground plant materials after oven drying at 65 °C for 48 h.
Soil samples were collected by taking three 7 cm diameter soil cores from 0 to 10 cm depth in each of the 1 × 1 m quadrats. Three cores from each quadrat were mixed in situ to form one composite sample. After removal of roots and gentle homogenization, the moist soil was sieved through 2 mm mesh and separated into two parts. One part was air dried to determine soil pH. The other was maintained fresh to measure SIN.
Soil pH was measured with an Accumet Excel XL60 pH meter (Fisher Scientific, Fair Lawn, USA) at a soil to water ratio of 1:2.5. SIN was extracted with 2 M KCl solution and the concentrations of NH4+–N and NO3−–N in the extracts were subsequently measured using a flow injection analyzer (FOSS, Höganäs, Sweden).
Data analyses
Meteorological data from 2001 to 2015 were obtained from 50 meteorology stations in Inner Mongolia and provided by the Chinese National Meteorological Information Centre. Daily air temperature and total precipitation for each site were interpolated using a geographic information system-based multiple-regression method. The partitioning of precipitation falling as rain versus snow was estimated using a simple temperature threshold (Berghuijs et al 2014). Specifically, precipitation on days with a daily average temperature above 1 °C was considered to be entirely rainfall, whereas on days with temperature below 1 °C, precipitation was considered to be entirely snowfall (Berghuijs et al 2014). The annual values used in this analysis are from 1 September to 31 August next year to minimize the effects of carry-over storage of snowfall. The community-weighted green-up date for each 1 × 1 m quadrat was calculated as the sum of each species' green-up date weighted by relative abundance in terms of density (Garnier et al 2004).
Linear mixed-effect models were used to evaluate the effects of snowfall augmentation treatments, sites and their interactions on the snow depth, species richness, soil pH, SIN, community-weighted green-up date, and AGB. In models, the snowfall augmentation treatments served as the fixed factors, and sites were included as the random factors. In addition, linear regression analyses were used to evaluate the relationships of snowfall augmentation, annual precipitation, annual rainfall, and annual snowfall with species richness, soil pH, SIN, community-weighted green-up date, and AGB. The effects were considered to be significantly different if P < 0.05. All statistical analyses were performed using SPSS 17.0 (SPSS Inc., USA).
Structural equation models (SEMs) analysis was performed to test the multivariate effects on species richness through hypothetical pathways of abiotic factors and biotic factors. To facilitate our analysis and interpretations, we classified variables into two explanatory groups before evaluating SEMs. These groups included (1) abiotic factors (soil pH and SIN) and (2) biotic factors (community-weighted green-up date and AGB). The statistical analyses were performed using Amos 21.0 (Amos Development Corporation, USA).
Results
Species richness
Snowfall augmentation significantly decreased species richness in the temperate steppe in Inner Mongolia, China (P < 0.001, figure 2(a), table S2). For each individual site, negative relationships between snowfall augmentation and species richness were observed in 90% of the sites, although the correlations were only statistically significant for four of the ten sites (all P < 0.05) and marginally significant for three of the ten sites (all P < 0.10) (figure 2(b), table S3). The strongest reduction in species richness induced by snowfall augmentation (i.e., highest slope absolute values) occurred in the low-rainfall sites (figure 2(c)). Species richness in the control plots increased linearly with annual precipitation across the ten sites at the regional scale (P = 0.009, figure 2(b)). We found poor correlation between the species richness–snowfall augmentation relationship and annual snowfall at the regional scale (P = 0.190, figure 2(d)).

Figure 2. Precipitation versus species richness. (a) Effects of snow depth on species richness for each site, and each color corresponds to one site. Values are means ± SE (n = 3). (b) Points indicate the mean species richness in the control plots versus annual precipitation at each site. The thick, dark-gray line is the spatial fit across the region's precipitation range under control conditions (y = 0.06x-11.05, R2 = 0.59, P = 0.009). The colored lines denote the within-site relationship between snowfall augmentation and species richness, and each color corresponds to one site. (c) The slope of the linear regression model at each site (species richness = a × precipitation + b) was positively correlated with annual rainfall at the regional scale (y = 0.0003x-0.17, R2 = 0.43, P = 0.039). (d) Poor correlation between the slope of the linear regression model at each site (species richness = a × precipitation + b) and annual snowfall at the regional scale (R2 = 0.20, P = 0.190). Therefore, the strongest reduction in species richness induced by snowfall augmentation occurred in the low-rainfall sites. However, the effects of snowfall augmentation on species richness did not show an obvious pattern along the site-level snowfall gradient.
Download figure:
Standard image High-resolution imageAbiotic and biotic factors
Snowfall augmentation significantly increased soil pH in the temperate steppe in Inner Mongolia, China (P < 0.001, figure 3(a), table S2). For each individual site, positive relationships between snowfall augmentation and soil pH were observed in all sites, although the correlations were statistically significant for seven of the ten sites (all P < 0.05) (figure 3(b), table S4). The strongest increment in soil pH induced by snowfall augmentation (i.e., highest slope absolute values) occurred in the low-rainfall sites (figure 3(c)). Soil pH in the control plots decreased linearly with annual precipitation across the ten sites at the regional scale (P = 0.003, figure 3(b)). In addition, snowfall augmentation significantly increased SIN (P = 0.008, figure S2, table S2).

Figure 3. Precipitation versus soil pH. (a) Effects of snow depth on soil pH for each site, and each color correspond to one site. Values are means ± SE (n = 3). (b) Points indicate the mean soil pH in the control plots versus annual precipitation at each site. The thick, dark-gray line is the spatial fit across the region's precipitation range under control conditions (y = −0.0033x + 8.49, R2 = 0.70, P = 0.003). The colored lines denote the within-site relationship between snowfall augmentation and soil pH, and each color corresponds to one site. (c) The slope of the linear regression model at each site (soil pH = a × precipitation + b) was negatively correlated with annual rainfall at the regional scale (y = −0.00004x + 0.02, R2 = 0.74, P = 0.001). (d) Negative correlation between the slope of the linear regression model at each site (soil pH = a × precipitation + b) and annual snowfall at the regional scale (y = −0.0006x + 0.01, R2 = 0.50, P = 0.023). Therefore, the strongest increment in soil pH induced by snowfall augmentation occurred in the low-rainfall and/or low-snowfall sites.
Download figure:
Standard image High-resolution imageSnowfall augmentation significantly advanced community-weighted green-up date in the temperate steppe in Inner Mongolia, China (P < 0.001, figure 4(a), table S2). For each individual site, negative relationships between snowfall augmentation and community-weighted green-up date were observed in 80% of the sites, although the correlations were only statistically significant for four of the ten sites (all P < 0.05) and marginally significant for two of the ten sites (all P < 0.10) (figure 4(b), table S5). Community-weighted green-up date in the control plots decreased linearly with annual precipitation across the ten sites at the regional scale (P = 0.025, figure 4(b)). In addition, snowfall augmentation significantly increased AGB (P = 0.005, figure S3, table S2).
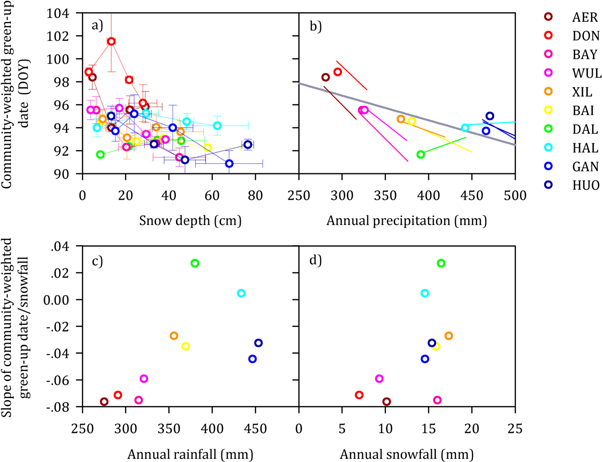
Figure 4. Precipitation versus community-weighted green-up date. (a) Effects of snow depth on community-weighted green-up date for each site, and each color correspond to one site. Values are means ± SE (n = 3). (b) Points indicate the mean community-weighted green-up date in the control plots versus annual precipitation at each site. The thick, dark-gray line is the spatial fit across the region's precipitation range under control conditions (y = −0.02x + 103.24, R2 = 0.49, P = 0.025). The colored lines denote the within-site relationship between snowfall augmentation and community-weighted green-up date, and each color corresponds to one site. (c) The slope of the linear regression model at each site (community-weighted green-up date = a × precipitation + b) was marginally correlated with annual rainfall at the regional scale (R2 = 0.39, P = 0.054). (d) Poor correlation between the slope of the linear regression model at each site (community-weighted green-up date = a × precipitation + b) and annual snowfall at the regional scale (R2 = 0.33, P = 0.080). Therefore, the effects of snowfall augmentation on community-weighted green-up date didn't show an obvious pattern along the site-level rainfall or snowfall gradient.
Download figure:
Standard image High-resolution imageTo quantify the relative importance of the different controls on species richness, we constructed SEMs analysis based on the relationships between species richness and its key drivers. Our model explained 60% of the variance of species richness (figure 5). The effect of snowfall on species richness through pathway of abiotic factor was −0.08 (0.28 × −0.28) for soil pH, whereas its effect through pathways of biotic factor was 0.02 (0.20 × 0.12) for AGB (figure 5). Similarly, the effect of rainfall on species richness through pathway of abiotic factor was 0.24 (−0.84 × −0.28) for soil pH, whereas its effect through pathway of biotic factor was not significant (figure 5).
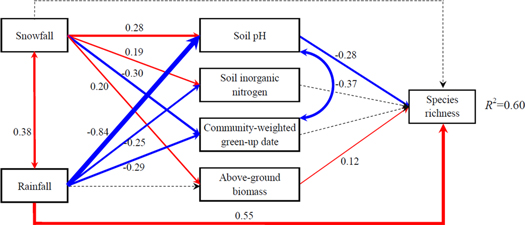
Figure 5. Structural equation models (SEMs) examining the effects of rainfall and snowfall on species richness via hypothetical pathways of abiotic factors and biotic factors. Solid red arrows represent positive paths (P < 0.05), solid blue arrows represent negative paths (P < 0.05), and dotted gray arrows represent non-significant paths (P > 0.05). Values associated with solid arrows represent standardized path coefficients, i.e., partial regression coefficients. Arrow width is proportional to the strength of the relationship. Also shown is the amount of variance explained (R2) in the species richness.
Download figure:
Standard image High-resolution imageSpecies richness decreased linearly with soil pH across the ten sites at the regional scale (P < 0.001, figure 6(a)). A negative relationship between species richness response and changes in soil pH was observed (P = 0.049, figure 6(b)).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 6. Relationships between soil pH and species richness across the ten snowfall augmentation experimental sites. (a) Species richness was negatively correlated with soil pH (y = −10.74x + 88.47, R2 = 0.50, P < 0.001). Each color corresponds to one site. (b) Response of species richness to snowfall augmentation was negatively correlated with response of soil pH to snowfall augmentation (y = −6.83x − 0.02, R2 = 0.40, P = 0.049). Therefore, the reduced species richness following 12 years of snowfall augmentation could have been attributed to the increased soil pH.
Download figure:
Standard image High-resolution image{kind=link}
Discussion
Increased snowfall decreases species richness rather than enhancing it
It has been proposed that increased precipitation notably enhanced species richness in arid and semiarid regions (Smith et al 1995, Zavaleta et al 2003, Stevens et al 2006). However, in our coordinated distributed experiments, snowfall augmentation decreased species richness rather than enhancing it (figures 2(a), (b)). Reduced species richness following 12 years of snowfall augmentation in this study could have been attributed to the increased soil pH (figures 5, 6(b)). A negative linear relationship between soil pH and plant diversity has been reported from global scale studies (Palpurina et al 2017) and regional surveys (Laliberte et al 2014). In addition, a slight positive linear relationship between snowfall augmentation and soil pH has been showed in the Gurbantunggut Desert, China (Fan et al 2013), and the McMurdo Dry Valleys, Antarctica (Ayres et al 2010). Our coordinated distributed experiments demonstrate the importance of soil pH for determining plant diversity under changing precipitation regimes in semiarid grasslands (Slessarev et al 2016).
Intriguingly, abiotic factor driven by soil pH was the dominant determinant affecting the variation in species richness under changing precipitation regimes, overriding biotic factor (figure 5). The prevailing view is that the mechanisms associated with biotic factor best explain the responses of species richness to global change (Tylianakis et al 2008). For example, species loss is expected as a consequence of increased productivity by upper vegetation, which exacerbates the limitation of available light to the lower canopy in a nitrogen enrichment experiment (Hautier et al 2009). The inconsistency of our result with the prevailing view may have been primarily due to the study scale. Biotic factor is especially relevant for explaining site-scale phenomena and testing community-scale hypotheses (Tylianakis et al 2008). However, site-specific information may not explain the variation in plant diversity among sites because of the community's variation (Fraser et al 2013). Conversely, abiotic factor is especially suitable for shaping plant diversity patterns at a regional scale, which has been reported in tropical rain forests (Jabot and Chave 2011), temperate grasslands (Bai et al 2007), and alpine tundra (Spasojevic et al 2013). Therefore, our coordinated distributed experiments suggest that the prevailing view of the mechanisms associated with biotic factor best explain patterns in plant diversity under climate change should be revisited.
Rainfall-dependent influence of increased snowfall on species loss
The strongest reductions of species richness induced by snowfall augmentation were found in the low-rainfall sites (figure 2(c)). Drier sites showed a higher snowfall sensitivity of plant diversity, while wetter sites showed a lower snowfall sensitivity of plant diversity, suggesting that the magnitude of species loss induced by snowfall augmentation was rainfall-dependent. Co-limitation theories state that responses of ecosystem processes to one resource will likely be regulated by another resource (Harpole et al 2011). For example, ecosystem carbon exchange responses to warming depended upon precipitation regimes in the temperate steppes (Xia et al 2009). Nitrogen enrichment positively affected soil micro-organisms only when water was sufficient (Zhang et al 2015). Here, our coordinated distributed experiments describe the first evidence that there is rainfall-dependent influence of snowfall on species loss.
Conclusions
Extreme precipitation (including rainfall and snowfall) in arid and semiarid regions are found to increase with climate warming (Peng et al 2010, O'Gorman 2014). Previous studies have found that increased snowfall increases the mean streamflow (Berghuijs et al 2014), favors plants with the C4 photosynthetic pathway (Weltzin and McPherson 2000), and increases net ecosystem exchange (Chimner et al 2010). However, the impact of increased snowfall on plant diversity in arid and semiarid regions remains relatively unknown. Our study provides the evidence that increased snowfall decreased species richness rather than enhancing it (opposite of patterns we observed for biomass production). There is rainfall-dependent influence of snowfall on species loss. Our study provides insights into the relationship between precipitation and biodiversity in arid and semiarid regions.
Acknowledgments
We acknowledge the students from Lanzhou University, Shanxi Agricultural University, and Hebei Agricultural University for their help with field work. This study was supported financially by the National Key Research and Development Program of China from MOST (2016YFB0501501 and 2017YFB0504000), the National Natural Science Foundation of China (31400393, 41401110 and 41671046).