Whereas humans can outrun horses over large distances (BBC News 2004),
because of their adaptation for endurance (Bramble and Lieberman 2004), their
swimming performance is mediocre compared to that of tuna and sailfish, and
flight is impossible. No wonder that the flight of animals and plants such as
birds, bats, insects and autorotating seeds has long since inspired mankind to
invent its own flying machines. Just over 100 years old, human-designed aircraft
have barely taken off on an evolutionary timescale. Recently engineers have
stepped up by designing small unmanned air vehicles at the scale of flying
animals and plant seeds that innovate by mimicking nature's successful design
principles for highly maneuverable and efficient flight. Here we feature current
biomechanics flight research and bioinspired design crème. By featuring the
work in nine papers of both fields side-by-side, and motivating the authors to
speculate how their work could inspire the other group, we hope to stimulate
future interactions between these adjacent fields of research. Here, we provide
an overview of the authors' research and designs accompanied by their
perspectives on the value of their work for the adjacent field.
The startling diversity of natural flight
Flight, including gliding and parachuting, evolved independently in many
lineages of organisms, from insects, dinosaurs, birds, reptiles and mammals to
plant seeds. Key evolutionary advantages of flight, such as energy efficient
habitat exploration and dispersal, have driven the evolution of a myriad of
wings that carry many members of these groups through the air. There is still
intense debate on how flight initially evolved: from the ground up or from the
canopy down. Regardless, flight must have started with a leap into thin air
during which aerodynamic force first became a strong selective force on the
evolution of organismal body plans. In keeping with this, our special issue
takes off with the gecko's amazing capability to direct its fall in mid-air.
How a tail can help make the most out of a leap into thin air
By studying geckos falling under the pull of Earth's gravity, Jusufi et
al (2010) elegantly combine observations on how geckos can right and turn
their body mid-air, with insightful mathematical models, and a robot gecko that
enables them to validate the principles of righting and turning, while falling
to Earth. The work of Jusufi et al (2010) suggests that inertial
appendages could simplify control of a variety of robots and unmanned aerial
vehicles, because this solution allows for simple body reorientation. An
airborne robot could maintain control authority at high angles of attack or zero
airspeed, where lift-based control mechanisms become less effective. Next-
generation, legged robots could swiftly navigate three-dimensional terrains
through periods of parachuting between arboreal and terrestrial supports. Such
robots could be stabilized by tails with multiple degrees of freedom to change
shape and instantaneous moment of inertia more efficiently, much like the tails
geckos have at their disposal (Higham and Russell 2010). This would enable these
robots to take a leap into thin air and land safely on all feet.

Figure 1. Robot gecko that can right itself midair during its fall to the ground. Image credit: Thomas Libby.
Slithering through the transition from parachuting to gliding
Socha et al (2010) investigate how aerial snakes extend their glide
trajectory down to the ground while slithering their body to adjust its shape.
They find that, regardless the height of fall provided to the gliding snakes in
their experiments, none of the snakes reaches an equilibrium glide path. This
illustrates how dynamically challenging the evolutionary transition from
falling, to parachuting, and gliding flight might have been. In their outlook,
Socha et al (2010) emphasize that a snake-inspired undulating robot that
is capable of slithering through air has not yet been developed. Development of
such a robot will most likely require challenging simulations and flow
visualization experiments with live animals to obtain a better handle on
snake-like aerial descent. Such unconventional descent approaches for climbing
snake-robots might well improve their descent performance, and could expand their
explorative capabilities.

Figure 2. Flying snake slithering through air. Image credit: Jake Socha.
Boosting glide performance through efficient exploitation of thermals
Ákos et al (2010) compare strategies of birds and unmanned air vehicles
to exploit thermals, hot columns of air rising in the atmosphere, to boost their
glide performance. Such soaring harvests potential energy from warmed rising air
that lifts both birds and aircraft. The challenge is finding the thermals in
time and staying in them long enough to maximize flight performance. These
workers show that bird and human pilot soaring strategies correspond and can be
successfully implemented in unmanned air vehicles. Ákos et al speculate
that even though bird feathers have no inherent sensory capabilities, they are
able to sense the airflow across the wing through the different mechanoreceptors
around the follicles of the feathers (Brown and Fedde 1993, Shim and Husbands
2007). It is possible that soaring birds that depend on the exploitation of the
atmospheric energy (thermal updrafts, wind gusts) can also detect local updraft
information along the entire surface of their wings from changes in the
distribution of pressure (Usherwood et al 2005) sensed by the
mechanoreceptors. Ákos et al argue that the design of an unmanned air
vehicle that could measure detailed airflow information similar to birds could
help these vehicles to fly along local updrafts and hence minimize energy loss.
Airflow sensing would also enable the unmanned air vehicles to find more easily
the center and the border of the thermal updraft.

Figure 3. Storks outfitted with GPS trackers that helped decipher their soaring strategies. Image credit: Zsuzsa Ákos.
From gliding to powered flapping flight
Tobalske (2010) shows how many of the smallest birds have specialized in being
either highly accomplished cruisers through bounding flight, such as
budgerigars, or effective hoverers, such as hummingbirds. He illustrates how
these different flight modalities have far-reaching implications for the body
plan, neural flight control, and aerodynamic mechanisms that these birds employ
to stay aloft. Based on an extensive overview of how small birds fly, Tobalske
(2010) suggests that micro-air vehicles should hover like a hummingbird and
cruise like a budgerigar. Hummingbirds are uniquely adept at hovering and flying
slowly, whereas intermittent flight appears to be an energy-saving strategy for
flying over a wide range of speeds. Mechanical and energetic power requirements
vary according to a U-shaped curve with flight speed, but the shape of this
curve, and efficiency of converting chemical energy into mechanical work, can
vary with wing design and kinematics (Ellington 1991, Thomas and Hedenström
1998, Tobalske et al 2003). To maximize efficiency over a broad range of
flight speeds, a useful solution would be to design a robot with the capacity to
alter wing posture and pause flapping intermittently as a function of flight
speed. The robot should hover like a hummingbird, using continuous flapping with
extended wings and long-axis rotation at the end of each half stroke. Then, like
a budgerigar, it should shift to flap-gliding at slow and moderate speeds and to
flap-bounding at fast speeds. A flexible wing design would enable it to
progressively flex its wings during upstroke to reduce drag as speed increases.
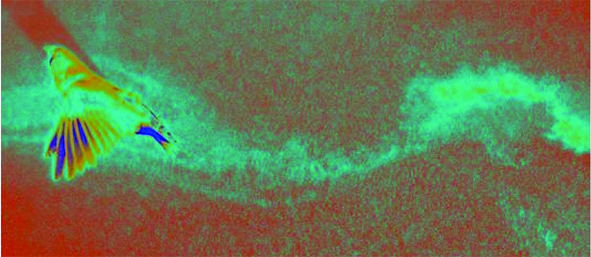
Figure 4. Airflow around a flying zebra finch seeded with a mist of micro olive-oil droplets. Image credit: Bret Tobalske.
Getting a grip on the physics of flight
Whereas biomechanics experiments are crucial for quantifying how organisms
fly, deeper insight into the underlying mechanisms can arise from studying the
physics of flight. In the past hundred years or so, our understanding of the
physics of airplanes has vastly increased our understanding of flight. One
remaining challenge, however, is that the aerodynamics of flight is highly
sensitive to the scale of organisms compared to that of aircraft, due to
so-called low Reynolds number effects. An even more challenging difference between
planes and organisms is the flexible morphology exhibited by flying animals.
Whereas the wings of maple seeds are similarly rigid as airplane wings, insect
wings are inherently flexible, bat wings have muscle fibers in their wing
membrane that can actively stretch the wing's surface, and birds can
dramatically change the shape of their wings through movement of joints and
overlapping feathers, while retaining an efficient aerodynamic shape. The
fluidity and flexibility of wing shape in these organisms are just starting to
be fully appreciated through physical analysis. We hope that this emerging field
will be further explored within a biologically relevant context. A promising
approach is the use of modeling techniques that can explore the parametric
design space of flexible wings, as presented below.
Flapping wing flexibility can be tuned for force control
Mountcastle and Daniel (2010) explore the aerodynamic performance of a
compliant flapping wing cross-section, the airfoil, with variable flapping
kinematics and variable chord-wise flexural stiffness. In their two-dimensional
model they combine an efficient vortexlet method with a basic finite element
method for flexible beams. The computational efficiency of this approach allows
for the quick exploration of the parametric design space of compliant wings. In
doing so, they find that lift and thrust of flexible wings are highly sensitive
to variations in chord-wise flexural stiffness, for which the performance optima
lie in different flapping-phase regions. This sensitivity could be employed by
animals, such as hawkmoths, to tune and control force production of their
flexible wings. Mountcastle and Daniel suggest that important flight performance
parameters such as lift and thrust of insect-scale wings can be tuned through
simple phase modulation. Whereas insects regularly actuate their wings along
three axes of rotation: sweep, elevation and pitch, their work suggests how
modulation between just two axes of actuation is sufficient and could simplify
control. A wide range of translational and rotational flight forces on the body
could be generated, simply via the control of sweep/pitch phase, employed
differentially between the left/right wing pair. Indeed, flies have been shown
to use a similar control strategy, generating yaw turning forces through
asymmetric timing of wing pitch rotation (Dickinson et al 1993).

Figure 5. Simulated vortex wake generated by a flexible airfoil (arrows indicate aerodynamic force). Image credit Andrew Mountcastle.
From complex to simple bioinspired airplanes
The burning question of how a growing understanding of the biomechanics of
flight can direct unmanned air vehicle design and inspire new conceptual
solutions to becoming and staying airborne remains largely unresolved. In our
opinion the main challenge is combining a deep insight of flight biomechanics
within its ecological, developmental and evolutionary context, with sound
engineering design principles based on a balanced mix of proven and novel
technology. Because biologists and engineers are typically trained quite
differently, which equips them with different scientific insights and
capabilities, this contributes to the gap between the biologist's understanding
of natural flight and the engineer's expertise in designing vehicles that
function well. In the middle are a few pioneering engineers who are able to
bridge both fields by mastering enough biological insight to design novel
biologically inspired air vehicles that work. Here we feature key designs that
not only work, but nicely illustrate how these designs can range from complex to
simple. These biologically inspired engineering solutions illustrate the
potential for bridging these two fields, but also demonstrate the current gap
between biology and current engineering designs. Whereas current engineering
designs benefit from simplicity, future ones might be more sophisticated with a
much wider performance envelope and broader range of applications inspired by
biology's vastly different scales of architectural organization and robust
multi-functionality. Below, we begin by considering complex biologically
inspired designs that fly and finish with successful simple designs that could
have a direct societal application now.
Gaining control of micro-flapping wings
Finio and Wood (2010) present several innovative insect-scale robotic thorax
designs capable of producing asymmetric wing kinematics similar to those
observed in nature and utilized by flies and other two-winged insects to
maneuver. Inspired by the thoracic mechanics of such insects, which entail a
morphological separation of power and control muscles, these designs show that
such distributed actuation can also modulate wing motion in a robotic design.
Although, Finio and Wood focus here on active control of wing kinematics for
body torque generation, which has been studied in insects, they also have a
parallel research area that focuses on passive body torque regulation using
purely mechanical feedback systems (Sreetharan and Wood 2010a, 2010b). For their
design work, it would be instrumental to know to what extent such passive
mechanical feedback is present in insect flight—for example, do insects
actively compensate for asymmetric wing loading due to perturbations, aging or
damage, or does some passive balancing mechanism allow the insect to continue
flight?

Figure 6. Thorax design of the Harvard robot fly. Image credit: Robert J Wood.
Mobile joints facilitate extreme wing morphing
Grant et al (2010) provide an overview of their micro-air vehicle
designs which are equipped with a series of mobile joints inspired by seagulls
to alter either the dihedral or sweep of the wings. This direct control over
extreme wing morphing enables these vehicles to trim with significantly
increased angles of attack and sideslip compared to traditional fixed-wing
aerial vehicles, particularly during a descent or in the presence of crosswinds.
Grant et al speculate that future designers will rely more and more on
experimental biology for a myriad of applications. These include the devices and
types of feedback used by nature to sense the flight environment, the
distribution of structural elements and actuation to maintain a desired shape
despite changes in loading, as well as the complex aerodynamics that result from
non-steady biological flight performance. These workers believe that our basic
understanding of the the brain could play a major role in design, as biologists
study information management and decision making in nature, which could provide
inspiration for novel autopilot design. Hence, a deeper understanding of how a
bird's brain controls flight would be much welcomed.

Figure 7. The mobile joints of seagull wings inspired Grant et al to design innovative morphing wings. Image credit: Mujahid Abdulrahim.
Losing your tail could be the next best thing
Hoey (2010) has constructed a series of radio-controlled glider models which
duplicate the aerodynamic shape of soaring ravens, turkey vultures, seagulls and
pelicans. Hence, his models have no vertical tail, although such a tail is
critical for the lateral stability of full-scale airplanes. Through flight
testing, Hoey determined the level of longitudinal and lateral-directional
static stability. Through a combination of basic theoretical flight dynamics and
clever experimentation, Hoey identified two factors that enable birds to be
passively stable while gliding without a vertical tail. First, the use of tail-
tilt to control small bank-angle changes, as observed in soaring birds, was
verified. Subsequent tests, using wing-tip ailerons, inferred that birds use a
three-dimensional flow pattern around the wing tip (wing tip vortices) which not
only generates a small amount of forward thrust in gliding flight, but also
controls adverse yaw to mediate lateral stability. Eliminating a vertical tail
has the potential of reducing both weight and drag of unmanned vehicles, and
could therefore find application in current designs. Hoey explains that there is
a significant unknown element as to what shape to model in his artificial birds,
since birds are highly flexible and are continually adapting the wing shape to
local conditions. Understanding how birds adjust their wing shape during gliding
and the extent to which this is under active versus passive control could be
extremely useful in furthering an understanding of how birds use their outer
wing panels for lateral stability and control.

Figure 8. These elegant models of a pelican, sea gull, and a turkey vulture can soar stable without a vertical tail. Image credit: Robert Hoey.
All you need is a single wing to take off
Ulrich et al (2010) present the first at-scale robotic maple seed.
Their design is directly inspired by their own observation of maple seed
geometries and descent dynamics when released from height. They found that body
roll and pitch angular rates for the various descent trajectories are coupled to
variations in wing pitch, which therefore provide a simple and direct means of
flight control. The novelty in the control strategy lies in its surprising
simplicity. Flight control through wing pitch not only allows for controlled
hovering and climb, but also lateral translation. This makes their vehicle not
only highly useful; it also shows how future micro-helicopters could be much
simplified compared to current designs. Ulrich et al find the flight
dynamics of their robot and maple seed to be substantially similar. They
speculate this could be of interest to those studying population dynamics of
samara-bearing trees as it provides a low-cost platform with a derived flight
dynamics model which can facilitate future testing of samara reaction to
horizontal winds. It also provides a platform for the testing of various wing
efficiencies, as the wing is interchangeable and can be used to measure the
power required to hover for a given geometry and flight modality, which may give
novel insight into the seeds' population dynamics.

Figure 9. At scale robotic maple seed. Image credit: Evan Ulrich.
Acknowledgements
We thank the authors for their exciting contributions to this special issue,
and their extra effort to provide a speculative outlook to further the adjacent
field; please refer to their paper first in case of making a direct reference to
one of the outlooks. We much appreciate the help and support of Andrew Malloy
and his editorial team to facilitate a high-quality peer-review process. DL is
supported by The Netherlands Organisation for Scientific Research—Earth and
Life Sciences Council (NWOALW grant 817.02.012). AAB is supported by NSF
IOS-074405 6.
References
Ákos Z, Nagy M, Leven S and Vicsek T 2010 Bioinsp. Biomim.5 045003
BBC News 2004 http://news.bbc.co.uk/2/hi/uk_news/wales/mid_/3801177.stm and Wikipedia http://en.wikipedia.org/wiki/Man_versus_Horse_Marathon
Bramble D M and Lieberman D E 2004 Endurance running and the evolution of Homo Nature432 345–52
Brown R E and Fedde M R 1993 Airflow sensors in the avian wing J. Exp. Biol.179 13–30
Dickinson M H, Lehmann F O and Gotz K G 1993 The active control of wing rotation by drosophila J. Exp. Biol.182 173–89
Ellington C P 1991 Limitations on animal flight performance J. Exp. Biol.160 71–91
Finio B M and Wood R J 2010 Bioinsp. Biomim.5 045006
Grant D T, Abdulrahim M and Lind R 2010 Bioinsp. Biomim.5 045007
Higham T E and Russell A P 2010 Flip, flop and fly: modulated motor control and highly variable movement patterns of autotomized gecko tails Biol. Lett.6 70–3
Hoey R G 2010 Bioinsp. Biomim.55 045008
Jusufi A, Kawano D T, Libby T and Full R J 2010 Bioinsp. Biomim.5 045001
Mountcastle A M and Daniel T L 2010 Bioinsp. Biomim.5 045005
Shim Y and Husbands P 2007 Feathered flyer: integrating morphological computation and sensory reflexes into a physically simulated flapping-wing robot for robust flight maneuver LNCS 4648 756–76
Socha J J, Miklasz K, Jafari F and Vlachos P P 2010 Bioinsp. Biomim.5 045002
Sreetharan P S and Wood R J 2010a Passive aerodynamic drag balancing in a flapping wing microrobotic insect Mech. Design132 051006–16
Sreetharan P S and Wood R J 2010b Passive torque regulation in an underactuated flapping wing robotic insect Robotics: Science and Systems (Zaragoza, Spain, June 2010)
Thomas A L R and Hedenström A 1998 The optimum flight speeds of animals J. Avian Biol.29 469–77
Tobalske B W, Hedrick T L, Dial K P and Biewener A A 2003 Comparative power curves in bird flight Nature421 363–6
Tobalske B W 2010 Bioinsp. Biomim.5 045004
Ulrich E R, Pines D J and Humbert J S 2010 Bioinsp. Biomim.5 045009
Usherwood J R, Hedrick T L, McGowan C P and Biewener A A 2005 Dynamic pressure maps for wings and tails of pigeons in slow, flapping flight, and their energetic implications J. Exp. Biol.208 355–69